Macaco-prego
Sapajus é um gênero de primatas da América do Sul que inclui as espécies de macaco-prego, também chamados micos-de-topete.[1] Os macacos do Novo Mundo do gênero Sapajus pertencem à família Cebidae, subfamíia Cebinae, e são uns dos mais comuns primatas da América do Sul. Outrora, as espécies desse gênero estavam incluídos no gênero Cebus e por muito tempo foram considerados membros de uma única espécie, Cebus apella. A taxonomia dos macacos-prego é confusa e mudou diversas vezes, sendo que o número de espécies variou entre uma única espécie até 12 espécies. Evoluíram, provavelmente, na região da Mata Atlântica e depois povoaram outros habitats, como a Amazônia. O registro fóssil é esparso. Ocorrem na América do Sul, desde regiões amazônicas, até o sul do Paraguai e norte da Argentina e por todo o Brasil. São muito adaptáveis e ocorrem virtualmente em todos os ambientes e tipos de vegetação ao longo de sua distribuição geográfica, desde florestas úmidas até savanas e o semiárido, como a Caatinga, do nível do mar até 2700 m de altitude.
| Macaco-prego | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Plioceno - Recente, 2,65–0 Ma | |||||||||||||||||||
 Quatro espécies de macacos-pregos, da esquerda para direita: Sapajus flavius, Sapajus xanthosternos, Sapajus nigritus, Sapajus libidinosus. | |||||||||||||||||||
| Classificação científica | |||||||||||||||||||
| |||||||||||||||||||
| Espécie-tipo | |||||||||||||||||||
| Simia apella Linnaeus, 1758 | |||||||||||||||||||
| Distribuição geográfica | |||||||||||||||||||
 Distribuição geográfica dos macacos-pregos.
S. apella S. cay S. flavius S. libidinosus S. macrocephalus S. nigritus S. robustus S. xanthosternos | |||||||||||||||||||
| Espécies | |||||||||||||||||||
| Sinónimos | |||||||||||||||||||
| |||||||||||||||||||
São macacos de porte médio, pesando entre 1,3 kg e 4,8 kg, com até 48 cm de comprimento, sem a cauda, que é preênsil, embora não tenha a mesma mobilidade que a cauda dos macacos-aranhas e muriquis. Esta tem papel principalmente na manutenção da postura. O quadrupedalismo é a forma predominante de locomoção. A cor da pelagem varia entre as espécies e os indivíduos, permitindo o reconhecimento destes. O pênis possui o formato de um prego quando ereto, o que conferiu o nome popular. Curiosamente, o clitóris também se assemelha a um pênis e dificulta a identificação dos sexos em indivíduos juvenis. O cérebro pesa cerca de 71 g e é complexo anatomicamente. O crânio e os dentes são robustos e apresentam adaptações à ingestão de alimentos duros e difíceis de mastigar.
Etimologia
editarO termo macaco-prego origina-se do fato de o pênis do animal ter o formato de um prego quando ereto.[2][3] O antigo gênero dos macacos-pregos, Cebus é uma palavra de origem grega, kêbos, e significa macaco com longa cauda.[4] A palavra Sapajus é um versão latinizada de uma palavra de origem tupi para os macacos-pregos.[5] Sapajou é a palavra francesa para macaco-prego.[5]
Taxonomia e evolução
editar

Os macacos-prego fazem parte da subfamília Cebinae, que está dentro da família Cebidae.[6] Essa subfamília é mais próxima, filogeneticamente, dos macacos-de-cheiro (subfamília Saimiriinae).[7] Elliot (1913) classificou as espécies do gênero Cebus, usando o táxon Simia capucina como espécie-tipo, definido por Linnaeus, em 1758.[6] Sapajus foi considerado como sinônimo de Cebus por Groves (2005), e foi um nome criado por Robert Kerr, em 1792, inicialmente como subgênero de Simia.[6][8] Para esse nome, a espécie-tipo é Simia apella, também definido por Linnaeus em 1758.[8] Von Pusch (1941) considerou Sapajus como subgênero de Cebus, e incluía as formas albifrons e apella.[8]
O gênero Cebus (gênero único na subfamília Cebinae) foi divido por Hershkovitz (1949) em dois grupos: os cebíneos com topete na cabeça e os que não possuem topete.[9] Essa separação também já foi feita por Elliot (1913), e foi defendida por outros pesquisadores após Hershkovitz.[10] Em 2012, Lynch Alfaro et al levaram essa divisão do gênero Cebus ao extremo, levando-se em consideração caracteres morfológicos e comportamentais, mantendo o gênero Cebus para os que possuem forma "grácil", sem topete e que habitam o bioma Amazônico, e separando o subgênero Sapajus em outro gênero, que inclui as espécies que possuem forma mais "robusta", topete e habitam as áreas de Floresta Atlântica, Cerrado e Caatinga.[8] Popularmente, as espécies de forma "grácil" e sem topete são conhecidas por "caiarara" e as que possuem forma "robusta" e topete, como "macaco-prego" (principalmente no Sudeste do Brasil).[11][12] Essa classificação foi defendida por Silva Jr em 2001.[8][11] Dados moleculares também sustentam essa divisão, onde os "cebíneos com topete" formam um clado e os "cebíneos sem topete" formam outro.[13][14]
A classificação das espécies de macacos-pregos é confusa e sofreu muitas alterações ao longo dos anos. Mesmo recentemente, muitos trabalhos divergem quanto ao número de espécies e subespécies, como observado por Rylands (2005) e Fragaszy & Visalberghi (2004).[10][15] Rylands (2000) atenta que a grande variabilidade individual dificulta a identificação de espécies e subespécies.[16] A dificuldade é maior ainda devido à hibridização entre as diversas formas, que ocorre devido a introdução de táxons originários de determinada região em outra, seja antes da colonização européia por indígenas, seja atualmente, por meio do comércio ilegal de animais silvestres.[16] Ao longo do século XX, o número de espécies variou significativamente, com Elliot (1913) considerando a existência de 12 espécies e com Hill (1960), considerando todas as formas como pertencentes a uma espécie, Cebus apella.[15] A partir da classificação de Groves (2001) e Rylands (2000), a espécie única C. apella de Hill foi dividida em quatro espécies diferentes.[15] Mas, antes disso, as espécies de macacos-prego da Mata Atlântica, como Sapajus xanthosternos e Sapajus nigritus já tinha sido propostas como espécies propriamente ditas por Torres (1983).[15] Silva Jr (2001), usando o conceito filogenético de espécie, considera algumas subespécies como espécies distintas, como é o caso da forma paraguayanus, de Sapajus libidinosus, classificada como Sapajus cay, a forma robustus de Sapajus nigritus e a forma macrocephalus de Sapajus apella.[11] Isso aumentou o número de espécies de macacos-prego para sete. A IUCN e outros autores, como Paglia et al (2012), consideram a classificação de Silva Jr (2001) como adequada.[12][17] Rylands fez uma revisão em 2005, mas utilizou a classificação de Groves (2001).[15] Em 2012, o trabalho de Ruiz-Garcia et al baseado em análises de DNA mitocondrial mostra um outro arranjo de espécies, mas considera que a classificação de Groves (2001) é mais provável que a de Silva Jr (2001).[18] Uma espécie, Sapajus flavius, foi recentemente redescoberta e considerada como espécie propriamente dita, somente 2006.[19][20]
| Hill (1960) | Rylands (2000) | Silva Jr (2001) | Groves (2001; 2005) |
|---|---|---|---|
|
|
|
Evolução e registro fóssil
editarAs duas formas de cebíneos evoluíram a partir do Mioceno (aproximadamente há 6,2 milhões de anos), quando um grupo ancestral ficou isolado na Amazônia e um grupo na Floresta Atlântica.[8][13] O gênero Sapajus surgiu desse grupo que ficou na Mata Atlântica, e a radiação das espécies neste bioma ocorreu há cerca de 2,65 milhões.[13] No Pleistoceno, há cerca de 400.000 anos, o gênero Sapajus foi para regiões do Cerrado e bacia Amazônica, com o surgimento de Sapajus cay.[13] Na Caatinga, Sapajus libidinosus surgiu há cerca de 200 mil anos.
O registro fóssil é esparso, e Chilecebus carrascoensis é um dos mais antigos fósseis próximos de Cebinae, encontrado no Chile.[21] Viveu há cerca de 20 milhões de anos, no Mioceno, e pesava cerca de 600 g.[21] Outras espécie, Killikaike blakei, descoberta em 2006, também parece se relacionar com os macacos-pregos e caiararas, e ocorreu na Argentina, no Mioceno, há cerca de 16,5 milhões de anos.[22] Essa espécie não possuía as mesmas características da face e dentição dos macacos-pregos modernos, mas já possuía significante capacidade cerebral.[22] A descoberta da espécie Acrecebus fraileyi na bacia do rio Solimões, sugere que Cebinae surgiu na Amazônia, há cerca de 9 milhões de anos, no Mioceno.[23]
Distribuição geográfica e habitat
editarOs macacos-pregos ocorrem no continente sul-americano, e duas espécies são endêmicas do Brasil (Sapajus xanthosternos e Sapajus flavius). São encontrados desde a bacia Amazônica até o sul do Paraguai e norte da Argentina e por todo o Brasil.[10][13] Por esses territórios, habita uma ampla variedade de formações vegetais, desde as florestas úmidas da Amazônia e Mata Atlântica, até áreas mais secas, como a Caatinga e o Cerrado, assim como o Pantanal, e o Chaco seco na Bolívia e Paraguai.[13][24]
_3.jpg)
Sapajus flavius e Sapajus xanthosternos são endêmicos do nordeste brasileiro. A primeira espécie ocorre somente no litoral dos estados de Pernambuco, Alagoas e Paraíba e sua ocorrência em regiões mais interiores, como na Caatinga, necessita confirmação, mas foi registrado uma população em áreas de Caatinga no Rio Grande do Norte.[19][20][25] S. xanthosternos ocorre no sul da Bahia e norte do estado de Minas Gerais, também em áreas de Mata Atlântica e de transição com a Caatinga.[10] Sapajus apella é uma espécie amazônica, ocorrendo no norte do Brasil, nos estados do Acre, Mato Grosso, Rondônia, Pará, Amapá e Roraima.[10] Também é encontrado na Amazônia no Peru, Equador, Colômbia, Venezuela, nas Guianas e em uma ilha isolada no litoral venezuelano, a ilha de Margarita.[10] Pela classificação de Silva Jr (2001), a espécie da Amazônia peruana, equatoriana e da Amazônia brasileira ocidental é S. macrocephalus.[13] Sapajus libidinosus possui a distribuição mais ampla, ocorrendo por todo o Brasil central, ao norte do rio Paraná e rio Grande e leste do rio São Francisco, em regiões mais secas dos estados de Minas Gerais, São Paulo, Mato Grosso do Sul, Mato Grosso, Goiás, Tocantins, Maranhão, Piauí, Ceará, Pernambuco, Rio Grande do Norte, Paraíba, Alagoas e Bahia.[10] Ocorre também no Paraguai e Bolívia.[10] Entretanto, pela classificação de Silva Jr (2001), o táxon encontrado no Paraguai e Mato Grosso do Sul corresponde a S. cay.[11] Sapajus nigritus é uma espécie típica do sudeste e leste brasileiro, sendo encontrado nos estados de São Paulo, Minas Gerais, Rio de Janeiro, Espírito Santo, Paraná, Santa Catarina e Rio Grande do Sul, e também no norte da Argentina.[10] Para Silva Jr (2001), que considera a forma robustus como uma espécie propriamente dita, a ocorrência de S. nigritus é limitada pela rio Doce ao norte, e entre esse rio e o Jequitinhonha, ocorre a espécie S. robustus.[10][11]

Deve-se salientar que os tipos predominantes de vegetação em determinada região da distribuição geográfica dos macacos-pregos são determinantes nos limites das ocorrências de algumas espécies de macacos-pregos.[26] Sapajus nigritus e S. robustus, apesar de terem como distribuição geográfica potencial áreas de Cerrado, são restritos a áreas de Mata Atlântica, principalmente em áreas úmidas do litoral.[26] Entretanto, em regiões de transição com a Mata Atlântica e Cerrado, é difícil determinar qual espécie ocorre, e no interior de São Paulo, no Brasil, não existe consenso se em algumas áreas ocorre Sapajus libidinosus ou S. nigritus.[26][27]
Os macacos-pregos são animais primariamente arborícolas, mas são muito adaptáveis e oportunistas, e podem viver em ambientes extremamente perturbados pelo homem, sendo encontrados, inclusive, em ambientes altamente industrializados e urbanizados.[10][24][28] Podem ser encontrados em qualquer tipo de floresta no Neotrópico, desde florestas deciduais e semideciduais, que passam entre 5 e 6 meses sem chuvas, até florestas pantanosas e manguezais.[10] Entretanto, já foi observado que S. apella evita áreas inundadas.[29] Ocorre em florestas ao nível do mar e em florestas montana e sub-montana até 2700 m de altitude, porém, parece preferir áreas de florestas de terras baixas.[10][24] Sua adaptabilidade permite que vivam em pequenos fragmentos de floresta secundária cercados por campos cultivados, florestas de galeria[29] e áreas de borda,[30] e são capazes de atravessar grandes distâncias com espaços abertos, como observado na Caatinga e no Cerrado.[10] Preferem os estratos médio da floresta, mas podem utilizar todos os estratos da floresta, inclusive o sub-bosque, para forragear, deslocar e beber água.[10][31] Por ora, os integrantes do gênero Sapajus mostram-se muito mais adaptáveis e oportunistas do que os caiararas, do gênero Cebus.[10][30] Visto a ampla distribuição geográfica dos macacos-pregos, os habitats e preferências de área de vida podem variar ao longo dos estudos realizados.[24]
Descrição
editar
Os macacos-pregos são macacos do Novo Mundo de porte médio, com os machos pesando entre 1,3 e 4,8 kg e as fêmeas entre 1,37 e 3,4 kg, tendo entre 35 e 48 cm de comprimento, sem a cauda, que possui entre 37,5 e 48,8 cm de comprimento.[32] Animais em cativeiro tendem a ser mais pesados que animais livres, e já foi registrado machos pesando até 6 kg.[3] Também, em cativeiro, podem viver até 55 anos, mas geralmente vivem até os 46 anos de idade.[33] Possuem um polegar pseudo-oponível na mão, pois a articulação carpometacarpal do polegar não é do tipo em sela.[3][24] Entretanto, isso não impede que o macaco-prego tenha precisão ao agarrar objetos pequenos, e de fato, é o único macaco do Novo Mundo que faz isso.[3]
A cauda é funcionalmente preênsil, entretanto, não possui todas as adaptações que a tornam verdadeiramente preênsil, como observado nos muriquis e macacos-aranhas, carecendo da região sem pelos na parte distal da cauda, que funciona como um coxim.[34] Quando em repouso, a cauda permanece totalmente enrolada, e pode ser utilizada como apoio, entretanto, ao contrário dos atelíneos ela não é utilizada para suportar o peso do corpo sozinha, e não funciona como um quinto membro, sendo curta, se comparada com muriquis, macacos-aranhas e bugios.[35][36] Ademais, a cauda é utilizada mais para manter a postura durante a alimentação do que para a locomoção, também contrastando com os muriquis e macacos-aranhas.[35] Rosenberger (1992) sugere que o surgimento da cauda preênsil dos macacos-pregos e caiararas deve-se ao relativo grande porte desses animais, contrastando uma dieta com significativa parcela de insetos, e por isso possui papel preponderante na manutenção de posturas durante o forrageamento e não para a locomoção.[37] Por conta do comprimento e flexibilidade menores, a cauda dos macacos-prego frequentemente é referida como "semi-preênsil".[3] O quadrupedalismo é a forma predominante de locomoção (cerca de 44%), mas podem saltar (cerca de 22%) e escalar quando se deslocam.[38][39] Nota-se, que quando comparado com o gênero Cebus, Sapajus se locomove de forma mais lenta, andando mais e correndo e pulando menos.[39] Ao contrário de outros macacos com cauda preênsil, não utilizam a braquiação para se locomover.[38] Quando se alimentam, costumam ficar sentados em substratos de tamanho médio.[38] A forma de se locomover e estratégias de forrageio determinaram a forma de outras estruturas do esqueleto pós-craniano dos macacos-pregos, além da cauda:os membros são relativamente curtos com relação ao corpo, e por isso os macacos-pregos possuem um aspecto atarracado.[8][39] Isto se relaciona com o fato de que dificilmente os macacos-pregos correm quando estão procurando alimento.[39] Quando comparado com Cebus olivaceus, possuem membros posteriores mais curtos, apesar de que não se observa tal diferença com os membros anteriores.[39] A escápula é relativamente alongada em relação a Cebus, o que sugere uma adaptação a escalar, apesar de que Sapajus não parece escalar significantemente mais do que Cebus.[38][39] Provavelmente, tal conformação se deve mais à manutenção de posturas sentadas ou bípedes no forrageio, como quando utilizam ferramentas.[39]

A coloração da pelagem varia, o que permite a identificação das espécies e dos indivíduos, mas todas as espécies possuem um visível tufo de pelos na cabeça, formando um topete, que não é evidente nos machos de Sapajus xanthosternos, embora nas fêmeas mais velhas dessa espécie, observa-se a presença deste.[8][10] Eventualmente, em S. apella macrocephalus o topete também é inexistente.[8] Quanto mais avançada é a idade, mais diferenciada é a pelagem entre os indivíduos, e mais fácil o reconhecimento destes e também, maior é o topete.[3] A cor do corpo varia desde um cinzento escuro ou preto em Sapajus nigritus e marrom a marrom escuro em Sapajus apella até um amarelo claro em Sapajus libidinosus.[10] Mas mesmo essa coloração pode variar muito dentro da espécie e em um mesmo grupo, e a identificação de uma espécie baseada em um único indivíduo de acordo com a pelagem pode ser problemática.[3] Os membros, a cauda e o topete tendem a ter uma cor mais escura, chegando ao preto, com exceção de Sapajus flavius, que possui o corpo inteiramente de cor dourada.[10][19] A cor da pelagem pode variar de acordo com a exposição ao sol, sendo que animais que se expões mais tendem a ter a pelagem mais escura.[3] A face não possui pelos no adulto.[3] A genitalia possui forma ambígua em ambos os sexos, podendo, inclusive, dificultar a diferenciação dos sexos em animais juvenis.[3] O pênis é relativamente longo e termina na glande que possui forma de disco, que quando ereto, confere o formato de prego, ao qual deu origem ao nome popular desse primata no Brasil.[3] O clitóris é desenvolvido e pode ser tão grande quanto um pênis, entretanto, ao contrário do que se observa nos atelíneos, ele não é pendular, e sim ereto, tendo um osso em sua estrutura (chamado clitoridis), da mesma forma que o pênis possui o báculo.[3][40] Nota-se que o clitóris possui um tamanho maior nos juvenis e recém-nascidos, e diminui de comprimento na idade adulta.[41] No trato gastrointestinal, observa-se que o ceco é muito reduzido, evidenciando a ingestão de alimentos relativamente fáceis de digerir e com alto valor energético.[3][42] O restante do sistema digestório não difere dos outros primatas, o que é consistente com uma dieta onívora.[42] Nota-se, também, que os macacos-pregos possuem grandes cérebros em comparação com o sistema digestório, sugerindo que uma tendência no aumento da encefalização e diminuição do sistema digestório ao longo da evolução desses primatas.[43]
O macaco-prego possui 54 cromossomos e a morfologia destes varia entre as espécies e populações, principalmente com relação a variações na heterocromatina constitutiva.[44] Sapajus xanthosternos possui o par de número 11 completamente distinto daquelas espécies classificadas anteriormente como Cebus apella.[45] Nestas últimas espécies, o cromossomo 11 apresenta um grande terminal de heterocromatina.[45] Entretanto, esse mesmo cromossomo 11 mostra um comprimento variável do trecho de heterocromatina entre as espécies e é ausente em Sapajus nigritus.[46] Esses trechos de heterocromatina possuem sequências repetitivas, nomeadas CapA e CapB.[44]
Sistema nervoso
editar.jpg)
.jpg)
O cérebro pesa cerca de 71 g e já foi constatado coeficientes de encefalização de cerca de 3,49, o que sugere grande capacidade cognitiva.[3][32][nota 3] O córtex cerebral possui sulcos e giros, permitindo, inclusive, a delimitação de lobos frontal, parietal, temporal, occipital e insular, com sulcos semelhantes àqueles do encéfalo humano: sulcos central, lateral e calcarino.[48] Isso contrasta com o cérebro de Callithrix jacchus e de outros saguis, que é liso e sem sulcos e giros.[48] Em contrapartida, citoarquitetonicamente, o córtex pré-frontal do macaco-prego é semelhante ao do macaco-rhesus.[47] De fato, essa região representa até 20% do volume cerebral e isso pode estar relacionado com o complexo repertório comportamental dos macacos-pregos.[49] As áreas visuais ocupam porção significativa da parte posterior do córtex cerebral, o que demonstra a importância da visão nesses animais.[50] Elas, também, são mais complexas quando comparadas com outros macacos americanos, e se assemelha ao do macaco-rhesus.[51] No córtex somatossensorial primário a representação das mãos e dedos ocupa porção significativa, além de que a porção relacionada à cauda também é expandida, embora não tanto quanto observado nos macacos-aranhas e muriquis.[3] A organização geral desse córtex sensorial também não é diferente de outros primatas, somente no fato de que a representação do tronco e membros aparece invertida em relação aos outros macacos.[52][53] No córtex motor primário, foi observado que existe certa lateralização e assimetria morfológica, de acordo com as mão que são preferencialmente utilizadas para realizar determinadas tarefas: o sulco central direito em animais canhotos tende a ser mais profundo do que aquele de animais destros.[54] Apesar de ter sido verificada tal lateralidade no uso das mãos, ela não é evidente e parece não surgir de forma espontânea, sendo observada somente quando os indivíduos realizam tarefas determinadas por humanos.[55]
Sistemas sensoriais
editarOs macacos-pregos discriminam cores, entretanto, como os outros macacos do Novo Mundo, os machos possuem uma visão dicromática, não distinguindo tons de laranja e vermelho, ao passo que algumas fêmeas possuem visão dicromática e outras, tricromática.[56] A acuidade visual e sensibilidade à luminosidade são semelhantes ao dos seres humanos e de outros macacos do Novo Mundo, mas a visão escotópica e sensibilidade temporal são um pouco melhores do que observado em seres humanos.[3] O olfato possui importância reduzida, como em outros primatas antropóides, mas existe evidências de que esse sentido é utilizado para encontrar alimento, e os macacos-pregos aprendem a reconhecer cheiro de frutos melhor do que carne de peixe.[57] Ademais, os macacos-pregos possuem o órgão vomeronasal.[3] Como animais com dieta frugívora, possuem notável sensibilidade à sacarose e pode identificar se um fruto é maduro por meio da concentração de açúcar.[3] De forma geral, esses primatas toleram e identificam grande variedade de sabores. A audição é também um sentido bastante utilizado pelos macacos-pregos, visto terem uma ampla variedade de sons para se comunicar.[3] Ela é tal forma desenvolvida, que os macacos-pregos podem discernir entre uma noz oca e uma maciça por meio de batidas, como seres humanos.[58] Provavelmente, como os outros macacos do Novo Mundo, esses animais escutam melhor na faixa entre 7 a 10 kHz e podem detectar sons cm até mais de 45 kHz.[3] O tato também é desenvolvido, e como outros primatas, os macacos-pregos possuem impressões digitais.[3] Esse sentido permite que esses primatas consigam discernir entre uma uva e uma pedra somente pelo toque das mãos.[3]
Morfologia dentária e do crânio
editarO crânio e a dentição dos macacos-pregos demonstra certa robustez quando comparado com outros macacos sul-americanos, principalmente com o gênero Cebus. O crânio tente a ser mais largo e a testa é projetada para frente, diferente de Cebus.[59] Apresenta, também, dimorfismo sexual acentuado, com os machos possuindo maiores graus de robustez, tendo, inclusive, uma crista sagital no topo do crânio.[59] Tal dimorfismo aparece nos primeiros estágios do desenvolvimento, e isso ocorre porque os machos crescem de forma mais rápida em determinado período do tempo.[60] O arco zigomático também é proeminente, resultado de um volumoso músculo masseter, assim como a crista sagital é resultado de um desenvolvido músculo temporal.[3][61] Tais músculos se inserem de forma mais anterior na mandíbula, se comparado com os outros macacos sul-americanos, o que amenta a força da mordida.[62] A mandíbula também é mais robusta se comparada com representantes do gênero Cebus sendo mais larga e resistente e apresenta dimorfismo sexual, sendo mais desenvolvida nos machos.[63] Tal conformação física é consequência de uma dieta adaptada a alimentos duros e difíceis de mastigar, como nozes e sementes (chamada de "durofagia").[59][61][63] Esses parâmetros morfológicos do crânio não variam somente entre os gêneros Sapajus e Cebus, mas também dentro do próprio gênero dos macacos-pregos: as subespécies de Sapajus nigritus, S. n. nigritus e S. n. robustus, possuem maior arcada dentária, prognatismo e volume craniano do que Sapajus libidinosus.[64][nota 4]
Como os outros Platyrrhini, os macacos-pregos possuem 36 dentes, com a seguinte fórmula dentária: , o que significa que cada hemi-mandíbula possui um canino, dois incisivos, três pré-molares e três molares.[65] Assim como o crânio, os dentes dos macacos-pregos demonstram adaptações à durofagia.[62] Comparado com Cebus, os caninos são mais arredondados e largos, assim como o esmalte dentário é mais grosso nos molares e pré-molares.[62][66] Estes, são relativamente largos, com amplas superfícies de oclusão,com cúspides baixas e arredondadas, próprias para quebrar itens alimentares duros, como em nozes e sementes.[66]

- Comparação entre crânios de Sapajus (macaco-prego) e Cebus (caiarara)
-
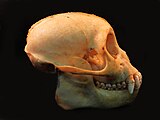 Crânio de Sapajus robustus, vista lateral.
Crânio de Sapajus robustus, vista lateral. -
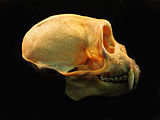 Crânio de Cebus olivaceus, vista lateral.
Crânio de Cebus olivaceus, vista lateral. -
 Crânio de Sapajus robustus, vista frontal.
Crânio de Sapajus robustus, vista frontal. -
 Crânio de Cebus olivaceus, vista frontal.
Crânio de Cebus olivaceus, vista frontal.




Ecologia e comportamento
editarOs macacos-pregos são animais diurnos e vivem em grupos, podendo ter até 40 indivíduos.[24] Esse número pode variar, dependendo do local, podendo ter número bem menores, de cerca de 5 indivíduos, em isoladas ilhas de floresta.[24] O território que tais grupos ocupam variam de tamanho, entre 81 e 293 hectares, e existe grande sobreposição entre os territórios de diferentes grupos.[67] A produtividade e distribuição de árvores frutíferas determina o tamanho dos territórios, existe uma correlação positiva entre o número de fêmeas do grupo e o tamanho da área de vida.[67] A interação entre os grupos pode ser pacífica, como observado em Manú, no Peru, ou agressiva, como em Iguazú,[68] na Argentina, o que evidencia mais uma das grandes variações observadas geograficamente entre os macacos-pregos.[67] O tamanho do grupo e relações dos indivíduos, assim como associações com outras espécies de primatas depende também da presença de predadores.[69]

Poucas observações de predação foram observadas para os macacos-pregos, e isso se deve provavelmente porque os próprios predadores evitam grupos observados por humanos, ou porque os locais de estudos incluem regiões em que os predadores já foram eliminados.[69] As taxas de predação também podem variar dependendo do local, e em Manu, no Peru, foi reportado que a cada duas semanas havia um episódio de predação, e no Parque Nacional Iguazú, na Argentina, foi observado isso em apenas duas ocasiões, após 750 dias.[70] Entretanto, observa-se que grandes aves de rapina como o gavião-real (Harpia harpyia) e gaviões do gênero Buteogallus provocam reações de defesas, como vocalizações de alarme e fuga.[24] Essas vocalizações podem ser emitidas mesmo para um jacaré.[24] Foram observados, além do gavião-real (que provavelmente é o maior predador dos macacos-pregos), a predação pelo uiraçu-falso (Morphnus guianensis).[71] Felinos, como a jaguatirica, a onça-pintada e a suçuarana, também são potenciais predadores de macacos-pregos e serpentes da família Boidae.[70][71] Também, macacos-pregos não habituados com seres humanos comportam-se como se estes fossem predadores.[69] Interessante notar, que na defesa antipredatória, os machos parecem ter um papel preponderante na vigilância e detecção de predadores, apesar de que todos no grupo mantêm-se atentos.[69]
Esses animais formam associações com outros primatas sul-americanos, como o macaco-de-cheiro (Saimiri), o bugio (Alouatta) e o macaco-aranha (Ateles).[24] Deve se salientar, que com os caiararas (Cebus), poucas formações de grupos mistos foram observadas.[24] Com os macacos-de-cheiro, as interações tendem a ser agressivas, com os macacos-pregos demonstrando dominância com relação aos primeiros.[24] Os macacos-pregos parecem se associar com os macacos-de-cheiro para aproveitarem a capacidade que esses últimos possuem de encontrarem insetos e outras presas. E apesar de terem sido reportadas associações de macacos-pregos com Ateles belzebuth, os primeiros evitam a interação com esses macacos-aranhas, principalmente em contextos competitivos.[24] Foi observado também, proximidade de grupos de macacos-pregos com primatas do gênero Chiropotes (os cuxiús) e do gênero Pithecia (os parauaçus, ou macacos-velhos), mas provavelmente isso se deu ao acaso, e não parece haver algum tipo de atração entre esses primatas.[24]
Escolhem altas árvores emergentes, com copas largas e ramos de grande diâmetro com forquilhas para dormir.[72] As fêmeas costumam dormir juntas e com sua prole, enquanto que machos periféricos dormem sozinhos em árvores próximas. A predação é o principal fator determinante na escolha dos sítios de dormida, e diferenças entre esses sítios ao longo da distribuição geográfica dos macacos-pregos parecem estar relacionadas com a estrutura da floresta e o padrão demográfico.[72]
Dieta e forrageamento
editarA anatomia, ecologia e distribuição geográfica dos macacos-pregos revelam uma dieta extremamente variável, sendo considerados animais onívoros, com adaptações à ingestão de alimentos duros ("durofagia").[8] Esse animais se alimentam desde itens de origem vegetal, até pequenos vertebrados.[67] São os mais "faunívoros" dos Platyrrhini e já foi registrado a predação de um filhote de guigó (Callicebus), o que sugere que eles podem predar outros primatas.[73] De fato, os macacos-pregos são os únicos macacos do Novo Mundo que se alimentam de outros mamíferos.[73][74] Também se alimentam de invertebrados aquáticos como caranguejos e ostras, o que não é observado, também, em nenhum outro macaco do Novo Mundo.[75][76] Os macacos-pregos também podem comer ovos de aves e provavelmente, de sapos.[24] A ingestão de alimentos de origem animal tem consequências no sistema digestório relativamente curto e em sua vida social complexa.[77] Porém, apesar desses fatos, a maior parte da dieta dos macacos-pregos é composta por frutos e insetos, e vertebrados correspondem entre 0,45 e 3,75% da dieta.[67][77]

Dentre as espécies de artrópodes consumidas, são conhecidos 17 táxons que são consumidos, principalmente insetos da ordem Hymenoptera e larvas de borboletas e mariposas.[77] É interessante que eles obtêm grande parte da proteína de origem animal através da insetivoria sem possuir grande especializações para esse tipo de dieta: eficientes técnicas de busca de insetos refletem na inteligência e habilidades manuais desses primatas.[77] O porte maior e robustez desses macacos, tal como suas habilidades, permitem que eles se alimentem de insetos maiores e que vivem escondidos, como no caso de formigas, ao contrário do que é observado com os macacos-de-cheiro.[77]
O número de espécies de plantas que são consumidas por macacos-pregos é grande, e consta com mais de 200 espécies.[67] Usualmente, os frutos são as principais porções ingeridas, entretanto, eles também comem flores e folhas.[24] Por conta de se alimentarem de frutos e sementes, existe um importante papel ecológico na dispersão de sementes.[24] As espécies de plantas que são importantes na dieta desses primatas variam de acordo com a região que habitam e da estação do ano. Em algumas áreas de florestas subtropicais da Argentina, onde ocorre significativa sazonalidade na disponibilidade de recursos alimentares, eles podem se alimentar predominantemente de folhas e bromélias.[78] Também na Argentina, mas em áreas mais úmidas, a dieta não difere muito da observada em regiões tipicamente tropicais.[78] No Parque Nacional de Manú, no Peru, as famílias Moraceae, Annonaceae e Palmae são as mais importantes no regime alimentar.[69] Em regiões altamente fragmentadas do sudeste do Brasil, macacos-pregos podem se alimentar de cultivos agrícolas humanos, como milho e cana-de-açúcar.[27] Durante a estação seca do ano, esses dois cultivos podem corresponder a cerca de 70% das plantas consumidas.[27] Apesar disso, mesmo me pequenos fragmentos cercados de campos cultivados, as plantas nativas possuem importante papel na dieta dos macacos-pregos.[27]
_Crab.jpg)
A ingestão de alimentos mais duros, como cocos e sementes, é um dos principais fatores na anatomia e comportamento dos macacos-pregos.[8] Em áreas secas, como a Caatinga no Brasil, é observado o uso de ferramentas para abrir a casca de cocos, o que não costuma ser observado em áreas mais úmidas, em que existe maior disponibilidade de frutos macios.[79][80] Essas ferramentas podem também ser utilizadas para se obter presas (artrópodes e pequenos vertebrados) e cavar[81][82][83]: esse comportamento permite acesso a alimentos antes inacessíveis, além de, em alguns casos, conferir economia no tempo do forrageamento.[79] É, de fato, um comportamento exclusivo entre os macacos do Novo Mundo.[79][80] O uso de ferramentas na natureza pelos macacos-pregos parece mais comum em regiões abertas, como a Caatinga e o Cerrado: apesar de observado manipulações complexas em macacos de outro biomas, como na Mata Atlântica e Amazônia, não caracterizam um uso de ferramentas propriamente dito.[80][84] Ademais, tal comportamento parece estar mais relacionado com hábitos de forrageamento terrestres, do que por pouca disponibilidade de alimento.[85][83]
Reprodução
editar
O estro é a cada 18 dias, e dura em torno de 5 ou 6 dias.[24][86] Em regiões com períodos de seca, existe sazonalidade no período de acasalamento, mas é possível observar cópulas durante o ano todo, com elas se concentrando na estação seca.[24][87] A gestação dura entre 5 e 6 meses (entre 155 e 162 dias), e as fêmeas geralmente têm um filhote por ano, mas pode ocorrer dois partos no mesmo ano.[24] Os nascimentos se concentram no início da estação chuvosa, entre dezembro e janeiro no hemisfério sul.[67]
Comportamento sexual
editarOs macacos-pregos possuem um repertório de corte e acasalamento dos mais complexos dentre os primatas não-humanos, e isso os diferencia dos caiararas (Cebus).[8] O sistema de acasalamento é considerado polígamo, com as fêmeas copulando com vários machos, e vice-versa.[24] Entretanto, há uma preferência da fêmea em copular com machos de posição elevada na hierarquia, e elas se engajam em comportamentos pró-ativos, solicitando a atenção do macho: exibições dos dentes e caretas, vocalizações e posturas submissas são exemplos de tal proceptividade.[86] Elas podem seguir o macho por até 4 dias, constantemente fazendo displays faciais, vocalizações específicas e até arremessando objetos para atrair a atenção do macho,[88] que demonstra indiferença e até mesmo as evita, e isto contrasta com o comportamento de outros machos de primatas.[86] Isso tem relação com os machos ejacularem apenas algumas vezes por dia e o macho alfa acaba sendo mais exigente com qual fêmea copula.[86] Isto não ocorre com machos subalternos, que precisam aproveitar as oportunidades de cópula.[86] Essa capacidade limitada de ejaculação, seja sazonalmente ou socialmente, é um fator importante na dinâmica sexual desses primatas.[24][87] Os macacos-pregos exibem comportamentos de corte mesmo após a cópula propriamente dita e a fêmea pode montar o macho, eventualmente.[8]
Conservação
editar
A maior ameaça aos macacos-pregos é a mesma a todas as espécies de primatas: a destruição do habitat e perturbação pelo homem reduz significativamente as populações das espécies de macacos-pregos.[10] A caça também é uma ameça e apesar de abundantes em muitas áreas, em outras, em que são caçados, como em regiões da Amazônia, as populações são significativamente reduzidas, e até extintos localmente.[24] Foi reportado, que vilas que sobrevivem com a caça e subsistência consomem até 200 macacos-pregos por ano, sendo superados apenas pelos grandes atelíneos.[89] Essa atividade também ameaça as populações de primatas do Nordeste Brasileiro, principalmente na Bahia.[90] Entretanto, a extrema adaptabilidade e flexibilidade com a dieta dos macacos-pregos permite que eles sobrevivam em áreas altamente industrializadas e fragmentadas, como em áreas de Mata Atlântica.[28] Em algumas localidades do estado de São Paulo, Espírito Santo e Minas Gerais, a densidade populacional é relativamente alta.[28]
Apesar de seu alto grau de conservação, a destruição da Amazônia é rápida e as espécies viventes em seus ambientes podem ficar em algum grau de ameaça futuramente. Entretanto, Sapajus apella[91] e S. macrocephalus[92] não correm grave risco de extinção e são considerados como com estado "pouco preocupante" pela IUCN. Entretanto, uma subespécie de S. apella, S. a. margaritae é considerada como "criticamente em perigo",[93] pois possui uma distribuição geográfica restrita, além das duas unidades de conservação em que ocorre sofrerem com o desmatamento ilegal.[94] S. macrocephalus é considerado como "quase ameaçado" no Equador, e foi extinto em algumas regiões desse país e do Peru, mas ainda existe em áreas remotas da bacia Amazônica e em unidades de conservação bem consolidadas.[94] S. apella e S. macrocephalus costumam ser caçados por subsistência em regiões que os grandes atelíneos já foram extintos, e também são perseguidos como pragas agrícolas, por conta de eventualmente invadirem plantações.[94]
Na Mata Atlântica, em que grande parte já foi desmatada, apenas Sapajus nigritus encontra-se como "quase ameaçada",[95] ao passo, que o macaco-prego-galego (S. flavius)[96] e o macaco-prego-do-peito-amarelo (S. xanthosternos)[97] encontram-se em estado "criticamente em perigo", segundo a IUCN, e já estiveram entre os 25 primatas mais ameaçados do mundo.[98] De fato, os macacos-pregos da região do Nordeste Brasileiro correm maior risco de extinção: até 2008, estimava-se que ocorriam apenas 180 indivíduos de S. flavius, mas estimativas mais recente apontam para uma população entre 1000 e 2000 indivíduos, em 26 fragmentos de Mata Atlântica, 15 deles no estado da Paraíba.[94] S. xanthosternos possui uma população de cerca de 3000 indivíduos, mas nenhuma subpopulação é viável a longo prazo.[94] O IBAMA não listou S. flavius em sua lista de espécies ameaçadas, mas também considera S. xanthosternos como "criticamente em perigo" e S. robustus[nota 5] como "vulnerável".[99] Essa última espécie, tem estado de conservação mais grave em Minas Gerais, e está incluído na lista de animais em extinção como "em perigo".[100] A IUCN também considera S. robustus como em "em perigo".[101] Para essa espécie, as unidades de conservação no Espírito Santo são preponderantes na preservação da espécie, por serem os maiores fragmentos de floresta em sua distribuição geográfica.[94]
As outras duas espécies que vivem fora da Mata Atlântica, em ambientes mais abertos, como o Pantanal e o Cerrado, Sapajus cay[102] e Sapajus libidinosus[103] não correm risco de extinção, e ambas são listadas como estado "pouco preocupante" pela IUCN. S. cay possui distribuição ampla, apesar de ser pouco conhecida, e ocorre em algumas unidades de conservação bem consolidadas no Brasil, Argentina e Paraguai.[94] S. libidinosus também possui distribuição ampla e é adaptável, mas é caçado onde ocorre e o desmatamento aumentou significativamente em seu habitat nos últimos 40 anos.[94]
| Espécie | População estimada | Estado na IUCN | Justificativa |
|---|---|---|---|
_2.jpg) Sapajus apella |
Desconhecido | Pouco preocupante (IUCN 3.1)[91] |
|
 Sapajus cay |
Desconhecido | Pouco preocupante (IUCN 3.1)[102] |
|
 Sapajus flavius |
1000-2000[94] | Criticamente em perigo (IUCN 3.1)[96] |
|
 Sapajus libidinosus |
Desconhecido | Pouco preocupante (IUCN 3.1)[103] |
|
_portrait.jpg) Sapajus macrocephalus |
Desconhecido | Pouco preocupante (IUCN 3.1)[92] |
|
.jpg) Sapajus nigritus |
Desconhecido | Quase ameaçada (IUCN 3.1)[95] |
|
.jpg) Sapajus robustus |
cerca de 8000[94] | Em perigo (IUCN 3.1)[101] |
|
 Sapajus xanthosternos |
cerca de 3000[94] | Criticamente em perigo (IUCN 3.1)[97] |
|
Notas de rodapé
editar- ↑ Nenhuma das classificações inclui Sapajus flavius pois ele foi considerado como espécie válida apenas em 2006.[19]
- ↑ O gênero Sapajus somente foi considerado como gênero propriamente dito a partir de 2012, por isso todas as classificações consideram o gênero Cebus.[8] Os autores se referem ao atual gênero Sapajus como as espécies do gênero Cebus que possuem topete na cabeça.[8] Silva Jr (2001) considera o termo Sapajus como subgênero.[11]
- ↑ Tal coeficiente é controverso, pois trata-se de um valor muito alto, mesmo quando comparado com chimpanzés, gorilas e orangotangos.[3]
- ↑ Neste trabalho, Masterson considerou todos os táxons como subespécies de Cebus apella.
- ↑ A lista do IBAMA considera a forma robustus de Sapajus nigritus como uma espécie separada (Cebus robustus).
Referências
- ↑ «Macaco-prego». Michaelis On-Line. Consultado em 21 de novembro de 2016
- ↑ «Macaco-prego». Dicionário Online de Português. Consultado em 2 de junho de 2013
- ↑ a b c d e f g h i j k l m n o p q r s t u v w x Fragaszy, D.M.; Visalberghi, E.; Fedigan, L.M. (2004). The Complete Capuchin: The Biology of Genus Cebus. Cambridge: Cambridge University Press. pp. 83–106. ISBN 0521667682
- ↑ William Rossiter (1879). An illustrated dictionary of scientific terms. London & Glasgow: William Collins, Sons, and Company. ISBN 0-548-93307-3
- ↑ a b «Sapajou». Collins American English Dictionary
- ↑ a b c d Groves, C.P. (2005). Wilson, D. E.; Reeder, D. M, eds. Mammal Species of the World 3.ª ed. Baltimore: Johns Hopkins University Press. pp. 136–138. ISBN 978-0-8018-8221-0. OCLC 62265494
- ↑ Chaterjee, H.J.; Ho, S.Y.; Barnes, I.; Groves, C. (2009). «Estimating the phylogeny and divergence times of primates using a supermatrix approach». BMC Evolutionary Biology. 9: 1-19. doi:10.1186/1471-2148-9-259
- ↑ a b c d e f g h i j k l m n o Lynch Alfaro, J.; Silva Jr, J. S.; Rylands, A. B. (2012). «How Different Are Robust and Gracile Capuchin Monkeys? An Argument for the Use of Sapajus and Cebus». American Journal of Primatology. 74 (4): 273-286. doi:10.1002/ajp.22007
- ↑ Hershkovitz, P. (1949). «Mammals of Northern Colombia, Preliminary Report No. 4: Monkeys (Primates), with Taxonomic Revisions of Some Forms». Proceedings of the United States National Museum. 98 (3232): 323-427
- ↑ a b c d e f g h i j k l m n o p q r s t u Fragaszy, D.M.; Visalberghi, E. (2004). «Taxonomy, distribution and conservation: where and what are they and how did they get there?». The Complete Capuchin: The Biology of the Genus Cebus. Cambridge: Cambridge University Press. pp. 13–36. ISBN 9780521667685
- ↑ a b c d e f Silva Jr., J. de S. (2001). Especiação nos macacos-prego e caiararas, gênero Cebus Erxleben, 1777 (Primates, Cebidae). Tese de Doutorado. Universidade Federal do Rio de Janeiro, Brasil
- ↑ a b Paglia, A.P.; da Fonseca, G.A.B.; Rylands, A.B.; Herrman, G.; Aguiar, L.M.S.; Chiarello, A.G.; Leite, Y.L.R.; Costa, L.P.; Siciliano, S.; Kierulff, M.C.M.; Mendes, S.L.; Tavares, V.C.; Mittermeier, R.A.; Patton, J.L. (2012). «Lista Anotada dos Mamíferos do Brasil 2ª edição» (PDF). Occasional Paper (6): 1-82. Consultado em 1 de junho de 2013. Arquivado do original (PDF) em 9 de novembro de 2013
- ↑ a b c d e f g Lynch Alfaro, J. W., Boubli, J. P., Olson, L. E., Di Fiore, A., Wilson, B., Gutiérrez‐Espeleta, G. A., Chiou, K.L.; Schulte, M.; Neitzel, S.; Ross, V.; Schwochow, D.; Nguyen, M. T.T.; Farias, I.; Janson, C. H. & Alfaro, M. E. (2012). «Explosive Pleistocene range expansion leads to widespread Amazonian sympatry between robust and gracile capuchin monkeys» (PDF). Journal of Biogeography. Journal of Biogeography. 39 (2): 272-288. doi:10.1111/j.1365-2699.2011.02609.x. Consultado em 31 de maio de 2013. Arquivado do original (PDF) em 26 de fevereiro de 2015
- ↑ Moreira, M. (2002). «SRY evolution in Cebidae (Platyrrhini:Primates)». Journal of Molecular Evolution. 55 (1): 92-103. ISSN 1432-1432. doi:10.1007/s00239-001-2308-7
- ↑ a b c d e f Rylands, A.B.; Kierulff, M. C.; Mittermeier, R. A. (2005). «Notes on the taxonomy and distributions of the tufted capuchin monkeys (Cebus, Cebidae) of South America» (PDF). Lundiana. 6 (supplement): 297-110. Consultado em 9 de junho de 2013. Arquivado do original (PDF) em 20 de junho de 2015
- ↑ a b Rylands, A.B. (2000). «An Assesment of the Diversity of New World Primates» (PDF). Neotropical Primates. 8 (2): 61-93. ISSN 1413-4703 Verifique
|issn=(ajuda) - ↑ «IUCN 2012». IUCN Red List of Threatened Species. Versão 2012.2. 2012. Consultado em 1 de junho de 2013
- ↑ Ruiz-García, M.; Castillo, M.I.; Lichilín-Ortiz N. (2012). «Molecular Relationships and Classification of Several Tufted Capuchin Lineages (Cebus apella, Cebus xanthosternos and Cebus nigritus, Cebidae), by Means of Mitochondrial Cytochrome Oxidase II Gene Sequences». Folia Primatologica. 83 (2): 100-125. doi:10.1159/000342832
- ↑ a b c d de Oliveira, M. M.; Langguth, A. (2006). «Rediscovery of Marcgrave's capuchin monkey and designation of a neotype for Simia flavia Schreber, 1774 (Primates, Cebidae).» (PDF). Boletim do Museu Nacional: Nova Série: Zoologia. 523: 1–16. Consultado em 1 de junho de 2013. Arquivado do original (PDF) em 5 de fevereiro de 2009
- ↑ a b Mendes Pontes, A. R.; Malta, A.; Asfora, P. H. (2006). «A new species of capuchin monkey, genus Cebus Erxleben (Cebidae, Primates): Found at the very brink of extinction in the Pernambuco Endemism Centre.» (PDF). Zootaxa. 1200: 1–12. Consultado em 4 de maio de 2009
- ↑ a b Flynn, J. J., Wyss, A. R., Charrier, R., & Swisher, C. C. (1995). «An early Miocene anthropoid skull from the Chilean Andes». Nature. 373 (6515): 603-607. doi:10.1038/373603a0
- ↑ a b Tejedor, M. F., Tauber, A. A., Rosenberger, A. L., Swisher, C. C., & Palacios, M. E. (2006). «New primate genus from the Miocene of Argentina». Proceedings of the National Academy of Sciences. 103 (14): 5437-5441. doi:10.1073/pnas.0506126103
- ↑ Kay RF & Cozzuol MA (2006). «New platyrrhine monkeys from the Solimões Formation (late Miocene, Acre State, Brazil)». Journal of Human Evolution. 50 (6): 673–686. PMID 16530809. doi:10.1016/j.jhevol.2006.01.002
- ↑ a b c d e f g h i j k l m n o p q r s t u v w Freese, C.H.; Oppenheimer, J.R. (1981). «The Capuchin Monkeys, Genus Cebus». In: Coimbra-Filho, A.F.; Mittermeier, R.A. Ecology and Behavior of Neotropical Primates - Volume 1. Rio de Janeiro: Academia Brasileira de Ciências. pp. 331–391
- ↑ Ferreira, R. G., Jerusalinsky, L., Silva, T. C. F., de Souza Fialho, M., de Araújo Roque, A., Fernandes, A., & Arruda, F. (2009). «On the occurrence of Cebus flavius (Schreber 1774) in the Caatinga, and the use of semi-arid environments by Cebus species in the Brazilian state of Rio Grande do Norte». Primates. 50 (4): 357-362. ISSN 1610-7365. doi:10.1007/s10329-009-0156-z
- ↑ a b c Vilanova, R.; Silva Jr, J.; Grelle, C.E.V.; Marroig, G.; Cerqueira, R. (2005). «Limites climáticos e vegetacionais das distribuições de Cebus nigritus e Cebus robutus (Cebinae, Platyrrhini)» (PDF). Neotropical Primates. 13 (1): 14-19
- ↑ a b c d Freitas, C. H. D., Setz, E. Z., Araújo, A. R., & Gobbi, N. (2008). «Agricultural crops in the diet of bearded capuchin monkeys, Cebus libidinosus Spix (Primates: Cebidae), in forest fragments in southeast Brazil». Revista Brasileira de Zoologia. 25 (1): 32-39. ISSN 0101-8175. doi:10.1590/S0101-81752008000100006
- ↑ a b c Pinto, N.; Lasky, J.; Bueno, R.; Keitt, T.H.; Galetti, M. (2009). «Primate Densities in the Atlantic Forest of Southeast Brazil: The Role of Habitat Quality and Anthropogenic Disturbance». In: Garber, P.; Estrada, A.; Bicca-Marques, J.C.; Heymann, E.W.; Strier, K. B. South American Primates: Comparative Perspectives in the Study of Behavior, Ecology and Conservation. New York: Springer. pp. 413–433. ISBN 978-0-387-78705-3. doi:10.1007/978-0-387-78705-3
- ↑ a b Defler, T. R. (1985). «Contiguous distribution of two species of cebus monkeys in El Tuparro National Park, Colombia». American Journal of Primatology. 8 (2): 101-112. doi:10.1002/ajp.1350080203
- ↑ a b Mittermeier, R. A., & Van Roosmalen, M. G. M. (1981). «Preliminary observations on habitat utilization and diet in eight Surinam monkeys». Folia Primatologica. 36 (1-2): 1-39. doi:10.1159/000156007
- ↑ Pontes, A. R. M. (1997). «Habitat partitioning among primates in Maraca island, Roraima, northern Brazilian Amazonia» (PDF). International Journal of Primatology. 18 (2): 131-157. ISSN 1573-8604. doi:10.1023/A:1026364417523
- ↑ a b Rowe, N. (1996). The Pictorial guide to Living Primates. Charlestown, Rhode Island: Pogonias Press. pp. 93–101. ISBN 0-9648825-0-7
- ↑ Fragaszy, D.M.; Visalberghi, E. (2004). «Life history and demography». The Complete Capuchin: The Biology of the Genus Cebus. Cambridge: Cambridge University Press. pp. 72–83. ISBN 9780521667685
- ↑ Ankel-Simons, F. «Survey of Living Anthropoidea». Primate Anatomy:An Introduction 3ª ed. Amsterdan: Elsevier. pp. 96–162. ISBN 978-0-12-372576-9
- ↑ a b Youlatos, D. (1999). «Tail use in capuchin monkeys» (PDF). Neotropical Primates. 7: 16-20
- ↑ Rosenberger, A. L. (1983). «Tale of tails: parallelism and prehensility» (PDF). American Journal of Physical Anthropology. 60 (1): 103-107. doi:10.1002/ajpa.1330600114
- ↑ Rosenberger, A. L. (1992). «Evolution of feeding niches in New World monkeys» (PDF). American Journal of Physical Anthropology. 88 (4): 525-562. doi:10.1002/ajpa.1330880408
- ↑ a b c d Youlatos, D. (1998). «Positional behavior of two sympatric guianan capuchin monkeys, the brown capuchin (Cebus apella) and the wedge-capped capuchin (Cebus olivaceus)» (PDF). Mammalia. 62 (3): 351-365. ISSN 0025-1461
- ↑ a b c d e f g Wright, K. A. (2007). «The relationship between locomotor behavior and limb morphology in brown (Cebus apella) and weeper (Cebus olivaceus) capuchins». American Journal of Primatology. 69 (7): 736-756. doi:10.1002/ajp.20391
- ↑ Carosi, M.; Haines, M. (1999). «The ocurrence of a bone-like structure in the clitoris of female tufted capuchins (Cebus apella)». American Journal of Primatology. 49 (1). 41 páginas. doi:10.1002/(SICI)1098-2345(1999)49:1<39::AID-AJP3>3.0.CO;2-9
- ↑ Carosi, M.; Gerald, M.; Ulland, A, Suomi, S. (2000). «Male-like External Genitalia in Female Tufted Capuchins (Cebus apella)». American Journal of Primatology. 51 (S1). 50 páginas. doi:10.1002/(SICI)1098-2345(2000)51:1+<33::AID-AJP3>3.0.CO;2-F
- ↑ a b Chivers, D.J.; Hladik, C.M. (1980). «Morphology of the gastrointestinal tract in primates: Comparisons with other mammals in relation to diet». Journal of Morphology. 166 (3): 337-386. doi:10.1002/jmor.1051660306
- ↑ Hartwig, W., Rosenberger, A. L., Norconk, M. A., & Owl, M. Y. (2011). «Relative brain size, gut size, and evolution in New World monkeys». The Anatomical Record. 294 (12): 2207-2221. doi:10.1002/ar.21515
- ↑ a b Seuánez, H.N.; Bonvicino, C.R.; Moreira, M.A.M. (2005). «The primates of the Neotropics: genomes and chromosomes». Cytogenetic and Genomic Research. 108: 38-46. doi:10.1159/000080800
- ↑ a b Seuánez, H.N.; Armada, J.L.A.; Freitas, L.; Rocha e Silva, R.; Pissinatti, A.; Coimbra-Filho, A.F (1986). «Intraspecific chromosome variation in Cebus apella (Cebidae, Platyrrhini). The chromosomes of the yellow breasted capuchin Cebus apella xanthosternos Wied, 1920.». American Journal of Primatology. 10 (3): 237-247. doi:10.1002/ajp.1350100304
- ↑ Ponsà1, M.; García, M.; Borell, A.; Garcia, F.; Egozcue, J.; Gorostiaga,M.A.; Delprat, A.; Mudry. M. (1995). «Heterochromatin and cytogenetic polymorphisms in Cebus apella (Cebidae, Platyrrhini)». American Journal of Primatology. 37 (4): 325-331. doi:10.1002/ajp.1350370407
- ↑ a b Roelf J Cruz-Rizzolo, Miguel AX De Lima, Edilson Ervolino, José A de Oliveira and Claudio A Casatti (2011). «Cyto-, myelo- and chemoarchitecture of the prefrontal cortex of the Cebus monkey». BMC Neuroscience. 12 (6). doi:10.1186/1471-2202-12-6
- ↑ a b Watanabe, L.S.; Madeira, M.C (1982). «The anatomy of the brain of the tufted capuchin (Cebus apella Linnaeus, 1758)» (PDF). Revista Brasileira de Odontologia. 11 (1-2): 5-12
- ↑ Borges, K. C. M., Ferreira, J. R., & Caixeta, L. F. (2010). «The prefrontal areas and cerebral hemispheres of the neotropical Cebus apella and their correlations with cognitive processes» (PDF). Dementia & Neuropsychologia. 4: 181-187
- ↑ Bonin, G.V. (1938). «The cerebral cortex of Cebus monkey». Journal of Comparative Neurology. 69 (2): 181-227. doi:10.1002/cne.900690203
- ↑ Rosa, M.G.; Piñon, M.C.; Gattass, R.; Sousa, A.P.B. «Third tier ventral extrastriate cortex in the New World monkey, Cebus apella». Experimental Brain Research. 132 (3): 287-305. ISSN 1432-1106. doi:10.1007/s002210000344
- ↑ Carlson, M.; Huerta, M.F.; Cusick, C.G.; Kaas, J.H. (1986). «Studies on the evolution of multiple somatosensory representations in primates: The organization of anterior parietal cortex in the new world callitrichid, Saguinus». Journal of Comparative Neurology. 246 (3): 409-426. doi:10.1002/cne.902460309
- ↑ Felleman, D.J.; Nelson, R.J.; Sur, M.; Kaas, J. (1983). «Representations of the body surface in areas 3b and 1 of postcentral parietal cortex of Cebus monkeys». Brain Research. 268 (1): 15-26. doi:10.1016/0006-8993(83)90386-4
- ↑ Phillips, K. A., & Sherwood, C. C. (2005). «Primary motor cortex asymmetry is correlated with handedness in capuchin monkeys (Cebus apella)» (PDF). Behavioral Neuroscience (6): 119. 1701-1704 páginas
- ↑ Fragaszy, D. M., & Mitchell, S. R. (1990). «Hand preference and performance on unimanual and bimanual tasks in capuchin monkeys (Cebus apella)». Journal of Comparative Psychology. 104 (3): 275-282. doi:10.1037/0735-7036.104.3.275
- ↑ Gomes, Ú. R., Pessoa, D., Tomaz, C., & Pessoa, V. F. (2002). «Color vision perception in the capuchin monkey (Cebus apella): a re-evaluation of procedures using Munsell papers». Behavioural Brain Research. 129 (1): 153-157. doi:10.1016/S0166-4328(01)00335-7
- ↑ Ueno, Y. (1994). «Olfactory discrimination of eight food flavors in the capuchin monkey (Cebus apella): comparison between fruity and fishy odors». Primates. 35 (3): 301-310. ISSN 1610-7365. doi:10.1007/BF02382727
- ↑ Visalberghi, E., & Néel, C. (2003). «Tufted capuchins (Cebus apella) use weight and sound to choose between full and empty nuts». Ecological Psychology. 15 (3): 215-228. doi:10.1207/S15326969ECO1503_2
- ↑ a b c Masterson, T. J. (1997). «Sexual dimorphism and interspecific cranial form in two capuchin species: Cebus albifrons and C. apella». American Journal of Physical Anthropology. 104 (4): 487-511. doi:10.1002/(SICI)1096-8644(199712)104:4<487::AID-AJPA5>3.0.CO;2-P
- ↑ Corner, B. D., & Richtsmeier, J. T. (1991). «Morphometric analysis of craniofacial growth in Cebus apella». American Journal of Physical Anthropology. 84 (3): 323-342. doi:10.1002/ajpa.1330840308
- ↑ a b Byron, C. D. (2009). «Cranial Suture Morphology and its Relationship to Diet in Cebus». Journal of Human Evolution. 57 (6): 649-655. doi:10.1016/j.jhevol.2008.11.006
- ↑ a b c Wright, B. W. (2005). «Craniodental biomechanics and dietary toughness in the genus Cebus». Journal of Human Evolution. 48 (5): 473-492. doi:10.1016/j.jhevol.2005.01.006
- ↑ a b Daegling, D. J. (1992). «Mandibular morphology and diet in the genus Cebus» (PDF). International Journal of Primatology. 13 (5): 545-570. ISSN 1573-8604. doi:10.1007/BF02547832
- ↑ Masterson, T. (2001). «Geographic Cranial Variation Among Three Subespecies of Cebus apella». American Journal of Primatology. 54 (S1): 46-47. doi:10.1002/ajp.1024
- ↑ Swindler, D.R. (2002). «Ceboidea». Primate Dentition An Introduction to the Teeth of Non-human Primates. Cambridge: Cambridge University Press. pp. 96–116. ISBN 978-0-511-06642-9
- ↑ a b Anapol, F., & Lee, S. (1994). «Morphological adaptation to diet in platyrrhine primates». American Journal of Physical Anthropology. 94 (2): 239-261. doi:10.1002/ajpa.1330940208
- ↑ a b c d e f g Fragaszy, D.M.; Visalberghi, E. (2004). «Behavioral Ecology: how do capuchins make a living?». The Complete Capuchin: The Biology of the Genus Cebus. Cambridge: Cambridge University Press. pp. 36–55. ISBN 9780521667685
- ↑ Di Bitetti, M. S. (2001). «Home‐range use by the tufted capuchin monkey (Cebus apella nigritus) in a subtropical rainforest of Argentina». Journal of Zoology. 253 (1): 33-45. doi:10.1017/S0952836901000048
- ↑ a b c d e Fragaszy, D.M.; Visalberghi, E. (2004). «Community ecology.How do capuchins interact with their local communities nd influence their environments?». The Complete Capuchin: The Biology of the Genus Cebus. Cambridge: Cambridge University Press. pp. 55–72. ISBN 9780521667685
- ↑ a b Hirsch, B. T. (2002). «Social monitoring and vigilance behavior in brown capuchin monkeys (Cebus apella)». Behavioral Ecology and Sociobiology. 52 (6): 458-464. ISSN 1432-0762. doi:10.1007/s00265-002-0536-5
- ↑ a b van Schaik, C.P.; van Noordwijk, M.A. (1989). «The special role of male Cebus monkeys in predation avoidance and its effect on group composition». Behavioral Ecology and Sociobiology. 24 (5): 265-276. ISSN 1432-0762. doi:10.1007/BF00290902
- ↑ a b Di Bitetti, M. S., Vidal, E. M. L., Baldovino, M. C., & Benesovsky, V. (2000). «Sleeping site preferences in tufted capuchin monkeys (Cebus apella nigritus)». American Journal of Primatology. 50 (4): 257-274. doi:10.1002/(SICI)1098-2345(200004)50:4<257::AID-AJP3>3.0.CO;2-J
- ↑ a b Sampaio, D. T., & Ferrari, S. F. (2005). «Predation of an infant titi monkey (Callicebus moloch) by a tufted capuchin (Cebus apella)». Folia Primatologica. 76 (2): 113-115. doi:10.1159/000083617
- ↑ Galetti, M. (1990). «Predation on the squirrel, Sciurus aestuans by capuchin monkeys, Cebus apella». Mammalia. 54 (1): 152-154. doi:10.1515/mamm.1990.54.1.147
- ↑ Port-Carvalho, M., Ferrari, S. F., & Magalhães, C. (2004). «Predation of crabs by tufted capuchins (Cebus apella) in eastern Amazonia». Folia Primatologica. 75 (3): 154-156. doi:10.1159/000078305
- ↑ Fernandes, M. E. (1991). «Tool use and predation of oysters (Crassostrea rhizophorae) by the tufted capuchin, Cebus apella appella, in brackish water mangrove swamp». Primates. 32 (4): 529-531. doi:10.1007/BF02381944
- ↑ a b c d e Janson, C. H., & Boinski, S. (1992). «Morphological and behavioral adaptations for foraging in generalist primates: the case of the cebines». American Journal of Physical Anthropology. 88 (4): 483-498. doi:10.1002/ajpa.1330880405
- ↑ a b Brown, A. D., & Zunino, G. E. (1990). «Dietary variability in Cebus apella in extreme habitats: evidence for adaptability». Folia Primatologica. 54 (3-4): 187-195. doi:10.1159/000156443
- ↑ a b c Moura, A. D. A., & Lee, P. C. (2004). «Capuchin stone tool use in Caatinga dry forest». Science. 306 (5703). 1909 páginas. doi:10.1126/science.1102558
- ↑ a b c Ottoni, E. B., & Izar, P. (2008). «Capuchin monkey tool use: overview and implications». Evolutionary Anthropology. 17 (4): 171-178. doi:10.1002/evan.20185
- ↑ Mannu, Massimo; Eduardo B. (1 de março de 2009). «The enhanced tool-kit of two groups of wild bearded capuchin monkeys in the Caatinga: tool making, associative use, and secondary tools». American Journal of Primatology (em inglês). 71 (3): 242-251. ISSN 1098-2345. doi:10.1002/ajp.20642
- ↑ Falótico, Tiago; Eduardo B. (1 de outubro de 2014). «Sexual bias in probe tool manufacture and use by wild bearded capuchin monkeys». Behavioural Processes. 108: 117-122. doi:10.1016/j.beproc.2014.09.036
- ↑ a b Falótico, Tiago; Siqueira, José O.; Ottoni, Eduardo B. (24 de julho de 2017). «Digging up food: excavation stone tool use by wild capuchin monkeys». Scientific Reports (em inglês). 7 (1). ISSN 2045-2322. PMC 5524703
 . PMID 28740211. doi:10.1038/s41598-017-06541-0
. PMID 28740211. doi:10.1038/s41598-017-06541-0
- ↑ Falótico, Tiago; Coutinho, Paulo Henrique M.; Bueno, Carolina Q.; Rufo, Henrique P.; Ottoni, Eduardo B. (17 de março de 2018). «Stone tool use by wild capuchin monkeys (Sapajus libidinosus) at Serra das Confusões National Park, Brazil». Primates (em inglês). 59 (4): 385–394. ISSN 0032-8332. doi:10.1007/s10329-018-0660-0
- ↑ Spagnoletti, Noemi; Visalberghi, Elisabetta; Verderane, Michele P.; Ottoni, Eduardo; Izar, Patricia; Fragaszy, Dorothy (2012). «Stone tool use in wild bearded capuchin monkeys, Cebus libidinosus. Is it a strategy to overcome food scarcity?». Animal Behaviour. 83 (5): 1285–1294. ISSN 0003-3472. doi:10.1016/j.anbehav.2012.03.002
- ↑ a b c d e Janson, C. H. (1984). «Female choice and mating system of the brown capuchin monkey Cebus apella (Primates: Cebidae)». Zeitschrift für Tierpsychologie. 65 (3): 177-200. doi:10.1111/j.1439-0310.1984.tb00098.x
- ↑ a b Alfaro, J. W. L. (2005). «Male mating strategies and reproductive constraints in a group of wild tufted capuchin monkeys (Cebus apella nigritus)» (PDF). American Journal of Primatology. 67 (3): 313-328. doi:10.1002/ajp.20188
- ↑ Falótico, Tiago; Eduardo B. (21 de novembro de 2013). «Stone Throwing as a Sexual Display in Wild Female Bearded Capuchin Monkeys, Sapajus libidinosus». PLoS ONE. 8 (11): e79535. PMID 24278147. doi:10.1371/journal.pone.0079535
- ↑ Chapman, C.A.; Peres, C.A. (2001). «Primate conservation in the new millennium: the role of scientists». Evolutionary Anthropology. 10 (1): 16-33. doi:10.1002/1520-6505(2001)10:1<16::AID-EVAN1010>3.0.CO;2-O
- ↑ Rylands, A. B., Mittermeier, R. A., & Rodriguez-Luna, E. (1997). «Conservation of neotropical primates: threatened species and an analysis of primate diversity by country and region». Folia Primatologica. 68 (3-5): 134-160. doi:10.1159/000157243
- ↑ a b Rylands, A.B., Boubli, J.-P., Mittermeier, R.A., Wallace, R.B. & Ceballos-Mago, N. (2008). Cebus apella (em inglês). IUCN 2013. Lista Vermelha de Espécies Ameaçadas da IUCN. 2013. Página visitada em 7 de julho de 2013..
- ↑ a b Rylands, A.B., Boubli, J.-P., Mittermeier, R.A., Stevenson, P., Palacios, E. & de la Torre, S (2008). Cebus macrocephalus (em inglês). IUCN 2013. Lista Vermelha de Espécies Ameaçadas da IUCN. 2013. Página visitada em 7 de julho de 2013..
- ↑ Ceballos, N. (2008). Cebus margaritae (em inglês). IUCN 2013. Lista Vermelha de Espécies Ameaçadas da IUCN. 2013. Página visitada em 7 de julho de 2013..
- ↑ a b c d e f g h i j k Anthony B. Rylands, Russell A. Mittermeier, Bruna M. Bezerra, Fernanda P. Paim & Helder L. Queiroz (2013). «Family Cebidae (Squirrel Monkeys and Capuchins)». In: Mittermeier, R.; Rylands, A.B.; Wilson, D. E. Handbook of the Mammals of the World - Volume 3. [S.l.]: Lynx. 952 páginas. ISBN 978-84-96553-89-7
- ↑ a b Kierulff, M.C.M., Mendes, S.L. & Rylands, A.B. (2008). Cebus nigritus (em inglês). IUCN 2013. Lista Vermelha de Espécies Ameaçadas da IUCN. 2013. Página visitada em 12 de julho de 2013..
- ↑ a b de Oliveira, M.M., Boubli, J.-P. & Kierulff, M.C.M. (2008). Cebus flavius (em inglês). IUCN 2013. Lista Vermelha de Espécies Ameaçadas da IUCN. 2013. Página visitada em 7 de julho de 2013..
- ↑ a b Kierulff, M.C.M., Mendes, S.L. & Rylands, A.B. (2008). Cebus xanthosternos (em inglês). IUCN 2013. Lista Vermelha de Espécies Ameaçadas da IUCN. 2013. Página visitada em 7 de julho de 2013..
- ↑ Mittermeier, R.A.;Schwitzer, C.; Rylands, A.B.; Taylor, L.A.; Chiozza, F.; Williamson, E.A.; Wallis, J. (2012). «Primates in Peril: The World's 25 Most Endangered Primates 2012–2014» (PDF). IUCN/SSC Primate Specialist Group (PSG), International Primatological Society (IPS), Conservation International (CI), and Bristol Conservation and Science Foundation (BCSF): 1–40. Consultado em 3 de junho de 2013. Arquivado do original (PDF) em 21 de outubro de 2013
- ↑ Chiarello, A.G.; Aguiar, L.M.S., Cerqueira, R.; de Melo, F.R.; Rodrigues, F.H.G.; da Silva, V.M. (2008). «Mamíferos». In: Machado, A.B.M.; Drummond, G.M.; Paglia, A.P. Livro Vermelho da Fauna Brasileira Ameaçada de Extinção - Volume 2 (PDF). Brasília, DF: Ministério do Meio Ambiente. pp. 680–883. ISBN 978-85-7738-102-9
- ↑ Vários autores (2007). «lista das espécies ameaçadas de extinção da fauna do estado de Minas Gerais». Biodiversitas. Consultado em 26 de junho de 2013
- ↑ a b Kierulff, M.C.M., Mendes, S.L. & Rylands, A.B. (2008). Cebus robustus (em inglês). IUCN 2013. Lista Vermelha de Espécies Ameaçadas da IUCN. 2013. Página visitada em 7 de julho de 2013..
- ↑ a b Wallace, R.B. (2008). Cebus cay (em inglês). IUCN 2013. Lista Vermelha de Espécies Ameaçadas da IUCN. 2013. Página visitada em 12 de julho de 2013..
- ↑ a b Rylands, A.B. & Kierulff, M.C.M. (2008). Cebus libidinosus (em inglês). IUCN 2013. Lista Vermelha de Espécies Ameaçadas da IUCN. 2013. Página visitada em 12 de julho de 2013..
Ligações externas
editar- ARKive – «imagens e vídeos do macaco-prego (gênero Cebus).» (em inglês)
- Animal Diversity Web – «Cebus apella tufted capuchin.» (em inglês)
- Folha de S.Paulo – «notícia sobre a redescoberta de Sapajus flavius, em 2006.»
- EthoCebus Project - «página sobre projeto de estudo do comportamento dos macacos-pregos.» (em inglês)