O grupo dos decápodes[1] inclui camarões, siris, lagostas, lagostins e caranguejos, sendo o grupo de crustáceos mais bem conhecido. São de tamanho grande, a maioria é comestível e têm importância econômica no que diz respeito à atividade pesqueira. Têm aproximadamente 15.000 espécies descritas,[2] constituindo um quarto de todos os crustáceos conhecidos e é ordem mais estudada. Seus representantes são de maioria bentônicos, mas há representantes de água doce (lagostins e algumas espécies de camarões), terrestres (algumas espécies de caranguejos) e planctônicos[3]
Para os taxóns inferiores consultar tópicos da página
A cabeça geralmente possui rostro fixo na posição mediana anterior, com dois pares de antenas birremes, exopodito de cada uma das segundas antenas com forma de escama antenal utilizada para controlar direção das correntes de água. A base das antenas contém o estatocisto e nefridióporo. Olhos compostos são pedunculados[3]
Os primeiros três segmentos torácicos são fundidos à cabeça para formar o cefalotórax. Os apêndice desses três segmentos são maxilípedes e atuam como peças bucais. Os outros cinco pares de apêndices torácicos são chamados de pereópodes, de onde deriva o nome Decapoda ( =10 pernas). O primeiro par de pereópodes forma um quelípode preênsil ou pinça, sendo mais desenvolvido e quelado. Geralmente os pereópodes não possuem exopoditos, portanto são unirremes. Epipoditos dos pereópode são modificados em brânquias.[3]
A carapaça se funde na região do dorso com os oitos segmentos torácicos. Os cinco últimos segmentos torácicos se distinguem do cefalotórax, mesmo estando cobertos e presos à carapaça. As projeções laterais da carapaça são mantidas afastadas do corpo e delimitam as câmaras branquiais laterais, onde se localizam as brânquias. O abdome, também chamado de pléon, é posterior ao tórax.[3]
Em 1951, a estimativa de espécies existentes de decápodos era de 8.31, distribuídas entre 1.000 gêneros, com Brachyura contabilizando 4.428 espécies.[4] Estudos mais recentes elevaram as estimativas de número de espécies para valores acima de 10.000 espécies.[5] Atualmente, acredita-se que existam 14.756 espécies de Decapoda, além de 3.300 fósseis, e 2.725 gêneros dentro do grupo, sendo Brachyura o maior deles, contendo 6.835 espécies.[6]
A atual classificação de Decapoda é baseada nos trabalhos iniciais de H. Milne Edwards, Dana, Latreille e seus contemporâneos. A partir de estudos mais sofisticados e análises moleculares e morfológicas foi possível chegar a uma classificação mais estruturada e estável, mas sem consenso total: [7]
Árvore filogenética dos grandes grupos de Decapoda, produzida baseada no artigo Phylogeny of Decapoda using two nuclear protein-coding genes: Origin and evolution of the Reptantia. Molecular Phylogenetics and Evolution.[8]
O tema central da evolução dos decápodes é a transformação de um ancestral alongado, natante e com corpo semelhante ao de um camarão, em um animal mais curto, achatado, reptante e morfologicamente semelhante a um caranguejo. A classificação dos decápodes é baseada na extensão dessa transformação. Apesar de a classificação dos decápodes ser muito complexa, para efeito de simplificação pode-se dizer que existem três formas de corpo básicas de decápodes: semelhante a um camarão (Penaeidea, Stenopodidea, Caridea), semelhantes a uma lagosta (Astacidea, Thalassinidea, Palinura) e semelhante a um caranguejo (Anomura, Brachyura).[3]
O corpo dos camarões tende a ser cilíndrico ou achatado lateralmente, com um abdome muscular bem desenvolvido. É comum observar a presença de um rostro serrilhado e em forma de quilha no cefalotórax desses organismos. As pernas são geralmente finas e os quelípodes podem estar presentes ou não. O exoesqueleto é comumente fino, flexível e não calcificado. Os pleópodes são grandes e apresentam franjas de cerdas, essas são as principais estruturas envolvidas na natação, embora haja mecanismos para a realização de movimentos bruscos para trás, como a rápida flexão ventral do abdome e o leque caudal, utilizados em situações de escape.[3]
Camarão- rosa, presente na costa leste dos Estados Unidos
A maioria dos camarões apresenta hábito bentônico e usam os pereópodes para andar sob o substrato. Embora seja possível nadar com o uso desses apêndices, esse tipo de deslocamento é intermitente nesses animais. Eles vivem entre as algas e outras macrófitas, embaixo de pedras e conchas e dentro de buracos e fendas de corais e rochas.[3]
Alguns, incluindo muito peneídeos e camarões do gênero Cragnon, cavam tocas rasas em substratos não consolidados por meio do batimento dos pleópodes.[9][10] Os padrões de atividade desses camarões podem estar relacionados com o fotoperíodo e com os ciclos de marés.[11] Já o camarão-rosaFarfantepenaeus duorarum, presenta na costa leste dos Estados Unidos, mantém-se quieto sobre o sedimento ou se enterra durante o dia, mas locomove-se ativamente durante o período noturno.[12] Nos Estados Unidos, a pesca do camarão concentra-se principalmente ao longo da costa sudeste do Atlântico e no Golfo do México. Os peneídeos representam o grupo mais primitivo dentro dos decápodes.[13]
Os camarões-de-estalo (Alpheidae) são comuns e apresentam distribuição bastante ampla.[14] Esses animais vivem em buracos e fendas, em esponjas ou embaixo de conchas, rochas e corais, ou ainda podem construir buracos ou abrigos.[15] Eles possuem de 3 a 6cm de comprimento e possuem uma das quelas muito maior que a outra. Há a presença de dois discos, um na base do dedo móvel erguido e outro no dedo fixo, que apresentam força adesiva entre si e evitam o fechamento espontâneo do dedo. Quando a contração muscular supera a força adesiva e o dedo móvel se fecha e nesse processo há a produção de um estalo devido ao processo de cavitação que ocorre próximo desses discos.[3][16] Esse mecanismo de estalo é utilizado para algumas espécies para a predação (quebrar pequenos mariscos e atordoar peixes), mas também é um mecanismo que funciona em confrontos agonístico (ameaça) entre indivíduos e provavelmente ajuda a manter um padrão uniforme de distribuição entre os membros da população.[17]
Indivíduo pertencente ao gênero Sergia, conhecidos por apresentarem bioluminescência
Alguns camarões de diversos táxons, conhecidos coletivamente como camarões limpadores, removem ectoparasitas e outros materiais indesejáveis das superfícies de peixes de recifes. O camarão pode subir em um peixe e até mesmo entrar na boca e na câmara branquial à procura de parasitas, os quais serão removidos e comidos.[3]
Camarões pelágicos que vivem a mais de 500 metros de profundidade (as zonas epipelágica e mesopelágica superior do oceano) tendem a ser transparentes ou semitransparentes e apresentam poucos cromatóforos laranjas e vermelhos , enquanto aqueles encontrados abaixo dos 500m apresentam uma coloração mais ou menos homogênea que varia do vermelho escuro ao vermelho escarlate.[18] Muitos desses últimos também apresentam órgãos bioluminescentes, ou fotóforos, localizados internamente ou na superfície do corpo, apresentando grande diversidade na morfologia dos fotóforos e em seu funcionamento e expressão.[19] Entre as espécies bioluminescentes mais bem conhecidas destacam-se as pertencentes aos aos gêneros Sergestes e Sergia cujo mecanismo de luminescência pode ser tanto por órgãos especializados quanto pela secreção na água ao redor do organismo formando uma espécie de nuvem luminosa[20]
Os outros integrantes de Decapoda são animais bentônicos adaptados para caminhar, semelhantes a lagostas ou caranguejos, e apresentando uma tendência do corpo ser achatado dorsoventralmente. As pernas, na maioria dos casos, são mais robustas que as dos camarões, com o primeiro par tipicamente adaptado em quelípodes. Os pleópodes não são adaptados para natação. Em muitos táxons, a forma primitiva do corpo encontra-se agora mais encurtada devido ao dobramento ventral do abdome sob o tórax .[3]
Upogebia major, representante dos Thalassinidea (decápodes de águas rasas ou entremarés).
Os decápodes semelhantes a lagostas, como as próprias lagostas, os lagostins e os talassinídeos, são formas intermediárias entre os caranguejos e os camarões.[3] Eles possuem um abdome similar ao dos camarões por ser grande e possuir todos os apêndices nessa região; além disso a carapaça costuma a ser mais comprida do que larga.[21] As lagostas e lagostins caminham usando os pereópodes, mas, de maneira semelhante ao camarão, podem escapar rápido para trás flexionando ventralmente o abdome e seu leque caudal.[22] Os pleópodes são usados para a ventilação, gerada a partir de uma corrente de água gerada pelo batimento desses apêndices que pode ser utilizado tanto para propósitos alimentícios quando respiratórios[23]
.
As lagostas são decápodes de corpo robusto e, em geral, habitam buracos e fendas em fundos rochosos ou coralinos. Membros dos Nephropidae possuem grandes quelópodes e são parecidos em sua morfologia aos lagostins (grupo próximo).[24] Um deles, a lagosta americana, Homarus americanus, pode atingir comprimento de cerca de 60cm e um peso entorno de 22kg. Essa espécie é pescada comercialmente com o uso de corvos ou armadilhas, nas quais o animal entra a procura de abrigo ou em busca de isca.[25] As caudas congeladas de lagostas vendidas em mercados são normalmente provenientes de Palinura,[26] procedente de diversas regiões tropicais e subtropicais do mundo, ocupando desde zonas litorâneas (rasas) até águas marinhas de profundidade (maiores que 400 metros).[27] Essas espécies não possuem quelípodes. Os talassinídeos (Calianassa, Thalassina e Upogebia) são decápodes de águas rasas ou da região entremarés, que vivem em galerias longas e profundas escadas na areia ou lodo.[28]
Representante da espécie Emerita brasiliensis, conhecido como tatúi, é muito ágil na locomoção sob a areia
Os decápodes remanescentes integram dois táxons, Anomura e Brachyura. Os Anomura representam a transição de camarões para caranguejos verdadeiros (os Brachyura). Sendo alguns representantes anomuros facilmente confundidos com caranguejos verdadeiros devido ao número de semelhanças, enquanto outros em nada se assemelham aos Brachyura.[29] Na maior parte das vezes, o abdome apresenta-se moderadamente reduzido, podendo ou não se apresentar flexionado sob o cefalotórax. Os urópodes estão presentes no abdome, com raras exceções. O quinto par de perna é reduzido e situa-se embaixo das laterais da carapaça ou dobrados sobre a porção posterior da carapaça.[30] A maioria dos anomuros é representada pelos ermitões. Esses decápodes singulares apropriam-se de conchas vazias de moluscos gastrópodes, usando-as como abrigos móveis[31] Na maioria desses organismos, o abdome é bem desenvolvido e não está flexionado sob o cefalotórax. Em vez disso, ele é assimétrico e adaptado para se encaixar dentro da câmara espiralada da concha que utilizam como abrigo.[32] O abdome vulnerável é recoberto por uma cutícula fina, flexível e não segmentada. No lado direito do abdome, os pleópodes são reduzidos ou estão ausentes, enquanto aqueles do lado esquerdo são mantidos nas fêmeas e usados para o carregamento dos ovos. A torção do abdome é adaptada para conchas dextrogira (espiraladas para a direita), embora as espiraladas para a esquerda também possam ser usadas. A maioria dos ermitões habita conchas de diferentes gastrópodes, dependendo da disponibilidade e oferta de conchas pode ocorrer maior ou menor número de trocas.[33][34] Nas tatuíras (Emerita brasiliensis), o modo característico de flexão do abdome e a forma esguia e ovoide do corpo representam algumas adaptações que permitiram que esse animal consiga se enterrar mais rápido na areia, iniciando pela região posterior do corpo. Os quelípodes estão ausentes nesses filtradores.[35] Os anomuros mais semelhantes aos caranguejos são os porcelanídeos. Esses organismos apresentam o abdome flexionado e simétrico, o que muito se parece com os caranguejos, mas não é tão reduzido quanto se faz em braquiúros. Os porcelaneídos são decápodes comuns de águas rasas em muitas partes do mundo, ocorrendo em grande número. Embora sejam muito semelhantes aos braquiúros, muitas espécies, quando incomodadas, realização atividade de natação mediante o batimento de flexões do abdome.[36] Os Brachyura possuem a forma de corpo mais especializada e, em termos de número de espécies representam os decápodes mais bem-sucedidos, totalizando cerca de 4.500 espécies conhecidas. Braquiúros podem ser encontrados na maioria dos habitats bentônicos marinhos, existindo ainda poucas espécies de água doce e terrestres. O corpo dele é fortemente deprimido e curto, tornando-o tão ou mais largo que longo. A carapaça cobre o cefalotórax e o péreon dorsal e lateralmente. Ventralmente, os segmentos do tórax são reconhecidos com facilidade, mas não são visíveis dorsalmente. O primeiro par de pereópode é, em geral, quelado e os demais são pernas com função locomotora ou, ocasionalmente, para a natação.[37]
Siri-azul (Callinectes sapidus) de grande importância econômica
O abdome é reduzido e firmemente flexionado sob o péreon, não sendo visível a não ser em vista ventral. Os urópodes estão ausentes, exceto em algumas espécies mais primitivas. Os caranguejos podem andar lentamente para frente, mas tipicamente se deslocam para os lados e com muita rapidez quando assim for necessário. Nesse modo de andar, as pernas-guia puxão mediante movimento de flexão, ao passo que as pernas acompanhantes empurram mediante movimento de extensão. Os quellípodes não são utilizados para andar.[3] A maioria dos caranguejos não pode nadar, mas os membros da família Portunidae, que inclui o comum e comestível siri-azul (Callinectes sapidus) da costa do Atlântico, são os mais poderosos e ágeis nadadores de todos os crustáceos[38] Nessa família, o último par de pernas de seus membros termina em largos remos achatados e durante a natação descrevem figuras de oito em seus movimentos. A ação é muito parecida à de um propulsor, e o quarto par de pernas age como um par de estabilizadores, uma vez que batem contrariamente[39] Os portunídeos podem nadar de lado, para trás e algumas vezes para frente com grande rapidez. No entanto, são animais bentônicos, como os outros caranguejos, e só nadam esporadicamente. Embora os quelípedes sejam importantes na defesa, outros dispositivos e hábitos protetores são observados na evolução dos braquiúros. Algumas espécies transportam anêmonas-do-mar em seus quelípedes.[40] Alguns caranguejos-aranha (apresentam corpos convexos triangulares e pernas delgadas) encontram-se cobertos com cerdas com ganchos, às quais se prendem em objetos estranhos. O caranguejo-decorador permanece relativamente imóvel e camuflado durante o dia, quando os predadores estão ativos.[3] Muitas espécies de caranguejos são capturadas e ingeridas por humanos em todo o mundo. O siri portunídeo Callinectes sapidus (o siri-azul) é o caranguejo comercial mais importante que ocorre ao longo das costas oriental e do Golfo dos Estados Unidos, especialmente na baía de Chesapeake.[41][42] Ele é capturado em águas rasas com uma armadilha ou linha, mas os pescadores comerciais também o capturam em grande número rebocando-os quando pescam camarões. Na costa ocidental dos Estados Unidos e na Europa, as espécies de Cancer (gênero de caranguejos não-nadadores) são utilizadas como alimento e são capturadas por armadilhas. Cancer magister (caranguejo-de-mercado) é a espécie mais importante da costa da Califórnia.[43]
A boca dos decápodes se localiza na superfície ventral da cabeça, ladeada e recoberta por diversas peças bucais, sendo que a região em torno da boca é chamada de quadro bucal, que é protegida por projeções, que não são apêndices, da parede do corpo. Seis pares de apêndices estão associados à boca dos representantes desse grupo.
O grupo dos decápodes apresenta uma ampla variação de hábitos e dietas alimentares, entretanto, na maioria das espécies associa alimentação predatória com saprofagia.[44] Existem representantes herbívoros, onívoros e detritívoros. A comida é agarrada com os quelípodes e passada para os terceiros maxílipes que a empurra para outras peças bucais. Enquanto o alimento é segurado pelas mandíbulas, as maxilas e maxilípedes retiram seus pedaços e os transferem para a boca.
Os quelípodes são adaptados às preferências e hábito de cada espécie. Decápodes que raspam algas de rochas ou se alimentam de detritos da superfície da areia e lodo, geralmente apresentam quelípodes com dedos no formato de colher. Os que se alimentam de moluscos possuem quelípodes dimórficos. A quela robusta e modificada para esmagar tem dentes rombudos, parecidos com molares, que são adaptados para quebrar conchas, ao passo que a quela cortante é mais delicada e adaptada para cortar carne.[45]
Detritívoros e suspensívoros se alimentam de partículas pequenas de alimento. Muitos decápodes são filtradores que passam a corrente de água através de uma malha de cerdas, que é utilizada para coletar partículas orgânicas em suspensão. Uma corrente própria de alimentação pode ser gerada pelos crustáceos, que também podem tirar vantagem de corrente já existente.[3]
O trato digestivo dos decápodes consiste em uma porção anterior elaborada e revestida por cutícula, de uma porção mediana de origem endodérmica e que possui cecos digestivos em desenvolvimento, e de outra porção posterior que também tem uma cutícula em seu revestimento. A função da porção anterior é trituração, hidrólise e separação de partículas pequenas que são digeríveis das que são grandes e não digeríveis. As partículas digeríveis e os solutos são encaminhados para os cecos digestivos, enquanto materiais que não podem ser aproveitados são expelidos pela boca ou então enviados para a porção mediana do trato digestivo, onde serão posteriormente eliminados.
Normalmente, o trato digestivo anterior tem um esôfago curto que é ligado a um estômago volumoso (proventrículo), que nos decápodes é dividido em uma grande câmara cardíaca anterior, e em outra câmara pilórica menor e posterior. Pelo fato de serem de origem ectodérmica, as duas câmaras são revestidas por um exoesqueletoquitinoso que possuem dentes e cerdas em suas paredes. Ambas as câmaras estão separadas uma da outra por uma constrição, pela qual há um fluxo nos dois sentidos. Geralmente, a região do dorso de cada câmara é especializada em tratar das partículas grandes e não digeríveis que serão encaminhadas para o intestino, enquanto as porções do ventre processam materiais orgânicos pequenos e digeríveis que são enviados aos cecos digestivos para absorção.
A câmara cardíaca tem um moinho gástrico revestido por filtro de cerdas e cutícula. O moinho gástrico é constituído por ossículos calcários (dentes) que maceram o alimento. A ação trituradora do moinho e o movimento das paredes do estômago são devidos à músculos extrínsecos presos ao ossículos.[46] Durante a trituração, a digestão que ocorre fora das células tem início com a mistura do alimento com enzimas que vêm dos cecos digestivos. O filtro de cerdas da câmara cardíaca apresenta cerdas com pouco espaçamento entre si, que acompanha o canal ventral que passa da câmara cardíaca para a pilórica. Depois da trituração e digestão, partículas finas e o soluto passam por meio do filtro de cerdas e entram a região ventral da câmara pilórica. As partículas grosseiras que não são digeridas e são rejeitadas pelo filtro de cerdas deixam a câmara cardíaca e atravessam a câmara pilórica pelo canal dorsal, e chegam até o intestino, local que ocorre a transformação em dejetos.
A câmara pilórica está dividida em um canal dorsal que leva diretamente até o intestino e uma complexa região ventral da qual os materiais digeríveis são encaminhados para os cecos digestivos. A região ventral consiste em um filtro mecânico e um filtro glandular. O alimento digerido e pré-filtrado pelo filtro de cerdas atinge uma região mediana muscular da câmara pilórica, chamada de filtro mecânico. O filtro glandular guarnece a abertura que leva aos cecos digestivos e é mais um outro filtro de cerdas com uma malha ainda menor que aquela do filtro de cerdas. O fluido é forçado por contrações dos músculos do filtro mecânico através do filtro glandular e para dentro dos cecos digestivos, enquanto o material rejeitado é encaminhado para o canal dorsal e para dentro do intestino. A prega pilórica, cujas cerdas evitam que partículas grandes entrem no filtro glandular, separa as porções dorsal e ventral da câmara pilórica.
Os cecos digestivos são órgãos grandes, de fundo cego, que liberam enzimas digestivas pelo epitélio de seus numerosos túbulos, que são enviadas para a câmara cardíaca do estômago.[47] Ocorre digestão intracelulare pinocitose de moléculas orgânicas solúveis, que são posteriormente armazenadas momentaneamente ou liberadas no sangue da hemocele.
As brânquias são compostas por um eixo central, correndo canais branquiais aferentes e eferentes, em que se arranjam extensões ou ramos laterais. O sangue de decápodos possui hemocianina dissolvida em seu plasma, e segue a partir do canal aferente para o interior dos respectivos filamentos, voltando então para o interior do canal eferente.[48] As respectivas estruturas para trocas gasosas surgem a partir da parede corporal ou próximo ao ponto de ligação do respectivo apêndice, recebendo diferentes nomes conforme sua localização.[49] O representante com maior número de brânquias é o camarãopeneídeoBenthesicymus, com 24, mas o comum é a redução desse número nos demais decápodos.
Para que ocorram efetivamente as trocas gasosas é necessária a produção de uma corrente ventilatória, através dos batimentos de um escafognatito ou alça branquial com forma de remo, projetado a partir da segunda maxila. A água é puxada para frente e a corrente flui para fora. Em camarões a água pode entrar na câmara branquial por qualquer ponto ao longo das bordas ventrais e posteriores da carapaça. Nos demais decápodos a carapaça geralmente se encaixa de maneira mais firme e assim a entrada da água se limita pelas margens posteriores da carapaça e ao redor das pernas.
Por poderem ser habitantes de fundo e também escavadores foram evoluídos mecanismos que evitam a obstrução das brânquias com resíduos ou sedimentos.[50] Nas bases dos quelípedes de caranguejos existem cerdas que filtram a corrente inalante, assim como nas coxas das pernas de lagostins e lagostas. Em alguns anomuros a limpeza das brânquias ocorre com o último par de pernas, enquanto que em alguns camarões pelos primeiros e segundos. Uma corrente reversa pode ser utilizada por algumas espécies escavadoras para a limpeza das partes ventrais do corpo, que podem estar cobertas por areia ou lama.
O coração está localizado no tórax e possui forma de caixa, apresentando três pares de óstios. Dele saem anteriormente cinco artérias e posteriormente uma artériaabdominal mediana, enquanto que do lado de baixo do coração surge uma artéria esternal. Cada uma dessas artérias principais é ramificada extensivamente e supre vários órgãos e estruturas.[49] O sangue é escoado no interior de um seio esternal mediano ventral antes de passar através das brânquias e retornar ao pericárdio e coração pelo trajeto de vasos branquiopericárdicos.[3]
Decápodos apresentam as glândulas das antenas (ou glândulas verdes), que são órgãos excretores, com o mais alto grau de desenvolvimento dentre os crustáceos.[49]São compostas pelo saco final (ou sáculo) e a primeira parte do túbulo (em forma de labirinto), localizados em frente e nos lados do esôfago. Por filtração o fluido é coletado dentro do sáculo, que possui podócitos nas paredes. As paredes do labirinto são bem pregueadas e glandulares, formando uma massa esponjosa e possibilitando uma área de reabsorção, ligando-se ao interior de uma bexiga por meio de um canal excretor. Partindo da bexiga se estende um pequeno duto no segmento basal da segunda antena, se abrindo para o exterior no ápice de uma pequena papila.
A maior parte dos resíduos nitrogenados são difundidos pela superfície corporal em que o exoesqueleto é mais fino, como por exemplo nas brânquias. As glândulas das antenas podem controlar a pressão do fluido interno e também as concentrações de íons específicos, como o Mg++. Entretanto, essas estruturas não exercem papel importante para a regulação osmótica.
Todos os caranguejos de água salobra e doce excretam uma urina que é isosmótica em relação ao sangue, e regulam o balanço iônico mediante a absorção de íons pelas brânquias.
Assim como em outros crustáceos, pernas e antenas exercem papel importante na detecção de estímulos ambientais. Antênulas possuem estetos, que localizam alimento e reconhecem outros membros da espécie.
Existem alguns decápodes cegos, em ambientes de grandes profundidades e entre os lagostins cavernícolas, os olhos composto pedunculados são geralmente bem desenvolvidos e o pedúnculo ocular apresenta mobilidade maior comparada com outros crustáceos. Os omatídios são compostos por córnea cuticular, cone cristalino tetrapartido, haste translúcida (somente em olhos que formam imagem por superposição), e uma retínula com oito células retinulares. Olhos de aposição e superposição estão presente nos decápodes.
Quase todos os decápodes possuem um estatocisto no artículo basal de cada antênula.[49] O estatocisto é uma cavidade preenchida com água, revestida por pequenas cerdas mecanorreceptoras. Os dendritos dos neurônios sensoriais ligados às cerdas levam ao cérebro a informação de deformações nas cerdas. Dentro da cavidade do estatocisto tem um estatólito, que repousa sobre as cerdas e pode sofrer deslocamento no interior do estatocisto, sob ação da gravidade. Essas estruturas se comunicam com o cérebro do animal pelo nervo antenular, o cérebro interpreta os sinais que chegam das cerdas receptoras para determinar a orientação do animal no campo gravitacional. Durante a ecdise alguns caranguejos perdem o senso de equilíbrio.
Cerdas mecanorreceptoras são receptores tônicos. Esses receptores enviam informações constantes de impulsos espontâneos ao cérebro, mesmo quando não há estímulos presentes.[51] Os receptores de fase, por outro lado, enviam ao cérebro um potencial de ação quando um evento ocorre e permanecem inativos em outros momentos.
O estatocisto e suas cerdas são estruturas do exoesqueleto que são substituídas, junto com o estatólito, a cada muda. O estatólito não é de origem epidérmica, portanto, não precisa ser reconstruído após a ecdise.
Caranguejos e alguns outros decápodes possuem estatocistos capazes de detectar aceleração em diferentes direções. Esses estatocistos possuem cerdas adicionais, distantes dos estatólitos, os quais são flexionadas pela inércia da água da cavidade do estatocisto. Por exemplo, em situação de repouso, a cutícula e água do interior do estatocisto estão estacionárias e os neurônios, na ausência de estímulos, enviam constantes sinais tônicos ao cérebro. Caso a cabeça se movimente, a cutícula do estatocisto acompanha o movimento na mesma velocidade que a cabeça, mas a água se movimenta depois. Isso faz com que as cerdas se movimentam através da água e acabam sendo flexionadas pela resistência inicial oferecida por ela. Os neurônios alteram a taxa de propagação dos sinais que são enviados ao cérebro, o qual interpreta isto como uma aceleração.
Os sistemas endócrinos e neurossecretor dos crustáceos e insetos são os mais bem conhecido dentre os invertebrados, contudo, o grupo dos decápodes foi o mais bem estudado. Eventos de duração prolongada ou contínua, como muda, reprodução e dispersão de cromatóforos, são controlados por hormônios. A maioria desses hormônios é produto de neurossecreção, liberados pelos axônios de neurônios secretores. Alguns órgãos endócrinos que são controlados pelo sistema nervoso também produzem alguns hormônios. Esses hormônios são liberados no sangue e são transportados até seus órgãos-alvo. O sistema neurossecretor mais importante dos decápodes se localiza no pedúnculo ocular, que possui olho composto em seu ápice, e dentro do pedúnculo, está o nervo óptico e vários conjuntos de células neuronais secretoras[52] Os hormônios sintetizados por células neurossecretoras do órgão X são liberadas pelos axônios para dentro da glândula do seio e depois para a hemocele. A circulação de sangue distribui esses hormônios até seus tecidos-alvo.
Os órgãos endócrinos conhecidos nos crustáceos são o órgão Y, os ovários, e a glândula androgênica. O órgão Y, que se localiza no cefalotórax, é controlado parcialmente por hormônios da glândula do seio e distribuído pela corrente sanguínea. Um dos hormônios secretados pelo órgão Y é a ecdisona, envolvido no processo de muda e é muito semelhante ao que está presente nos insetos.
Cromatóforos são células caracterizadas por armazenarem pigmentos, normalmente estão localizadas no tecido conjuntivo abaixo da epiderme e em áreas em que o exoesqueleto é fino ou transparente, contribuindo para conferir coloração aos crustáceos. A cor exibida por cada cromatóforo é variável e sua função é mudar a cor do animal visando a termorregulação ou a camuflagem. Um mecanismo pelo qual os cromatóforos podem mudar a cor do animal é por meio de divisões ao longo de um período de muitos dias, aumentando, dessa maneira, o número de cromatóforos e a quantidade total de pigmentos. De maior interesse, entretanto, é a capacidade dos cromatóforos de mudar de cor rapidamente e de maneira reversível, sem envolver alterações no número de células ou na quantidade de pigmento presente.
O cromatóforo dos crustáceos é uma célula com grânulos de pigmento intracelulares e com processo citoplasmáticos ramificados. As células ocorrem em grupos, mas geralmente como unidades independentes, formando órgãos cromatofóricos multicelulares com cooperação entre as células. Os grânulos de pigmento migram do interior de cada célula para alterar o impacto visual do órgão inteiro. Quando os grânulos disponíveis estão dispersos e ocupando o máximo da superfície, seu impacto visual é o maior possível. Quando eles estão concentrados na forma de uma pequena mancha no centro do órgão, o impacto é minimizado.[53]
São conhecidos cromatóforos com pigmentos branco, vermelho, amarelo, azul, marrom e preto. Os pigmentos vermelhos, amarelos e azuis são derivados de caroteno obtido na própria alimentação. Já a cor avermelhada tão marcante em caranguejos, lagostas e camarões é resultado de um carotenoide. No exoesqueleto dos animais vivos, esse carotenoide está conjugado com uma proteína e apresenta coloração azul. A fervura desnatura a proteína e altera a cor para vermelho. Por fim, pigmentos marrons e pretos são frutos das melaninas. Em alguns camarões, um mesmo cromatóforo pode ter um ou mais pigmentos diferentes, os quais podem migrar independentemente uns dos outros. Quase sempre, entretanto, a maioria dos cromatóforos é monocromática.
O tipo mais comum de mudança rápida de cor é o simples clareamento ou escurecimento característico de muitos caranguejos,[54] incluindo o chama-maré Uca, os quais são mais escuros durante o dia e mais claros durante a noite. O ritmo é endógeno e persiste na ausência de estímulos ambientais. Entretanto, há casos de crustáceos que são capazes de mudança de cores mais elaboradas, apresentando uma ampla variação de tonalidades. O movimento dos pigmentos nos cromatóforos é controlado por hormônios provenientes da glândula do seio.[55]
A migração dos pigmentos nos cromatóforos, nos omatídeos e outros processo fisiológicos frequentemente evidencia uma atividade rítmica nos crustáceos. Esses ritmos são controlados por relógios biológicos e têm sido estudados em várias espécies, em particular, no caranguejoCarcinus meanas, e no caranguejo chama-maré, Uca.[54]Ambos vivem em superfícies planas arenosas e na zona entremarés de praias protegidas, mas Carcinus é ativo durante a maré alta enquanto Uca é ativo quando o substrato fica exposto durante a maré baixa. Mediante a dispersão e concentração de pigmentos dos cromatóforos, ambas as espécies se tornam mais claras à noite mais escuras durante o dia. O ritmo endógeno persiste quando os caranguejos são mantidos constantemente em claro ou escuro. A remoção dos pedúnculos oculares, os quais secretam o hormônio de controle dos pigmentos, interrompe o ritmo em Carcinus e reduz sua amplitude em Uca.[55]
A atividade locomotora em ambos os caranguejos segue a variação da maré e é regulado por um relógio biológico relacionado ao ciclo lunar. Após cerca de uma semana sob condições constantes, o ritmo é perdido. Ele pode ser restaurado pela simples imersão de Uca em água do mar ou submetendo Carcinus a temperaturas mais baixas.
Ritmos semelhantes estão sendo observados em outras espécies de crustáceos, como o siri azul (Callinectes sapidus) e a baratinha-da-praia (Ligia excotica).
A maioria dos decápodes é dioica, embora alguns sejam hermafroditas.[56] A transferência de espermatozoides é indireta e acontece a partir da produção de espermatóforos, envolvendo comumente a cópula. Já a fertilização pode ser tanto interna quanto externa.
Em geral, os machos decápodes possuem um par de testículos que estão conectados aos gonóporos por um par de dutos espermáticos. Os gonóporos estão localizados no oitavo segmento torácico. Já os testículos estão localizados em posição dorsal no tórax e/ou abdome e apresentam variedade morfológica dentro dos grupos. O grau de desenvolvimento do duto espermático também varia em função, principalmente, da extensão de envolvimento desse na formação do espermatóforo. Os espermatozoides não possuem uma parte mediana nem flagelo, como o humano, e apresentam forma que lembra uma tachinha ou estrela.
É possível encontrar organismos nesse grupo que apresentam pênis, mas não são usados como estrutura introdutória. Ao invés disso, os pleópodes mais anteriores (primeiros segmentos abdominais), chamados de gonópodes, são modificados para atuarem como estruturas para a transferência indireta dos espermatóforos para a fêmea. Os pênis, com os gonóporos em suas extremidades, normalmente, transferem os espermatozoides dos testículos para os gonópodes ou, em situações mais raras, diretamente para a fêmea (não sendo esse caso o mais comum). Por exemplo, muitos branquiúros apresentam o primeiro pleópode em forma de um tubo longo, fino e aberto em ambas as extremidades; enquanto o segundo pleópode é longo, semelhante a um êmbolo fino que se encaixa dentro desse tubo. O pênis, curto e flexível nesse grupo, deposita o espermatóforo na base do tubo e o êmbolo desliza dentro do tubo, transferindo, assim, o espermatóforo para a fêmea.[3]
As fêmeas quase sempre possuem um par de ovários, localizados dorsalmente no tórax e abdome e conectados ao exterior por um par de ovidutos. Os ovidutos abrem-se ventralmente no sexto segmento torácico. Na condição mais primitiva, os espermatóforos são simplesmente grudados no corpo da fêmea, mas na maioria dos decápodes eles são injetados para dentro de um receptáculo seminal interno ou externo, no qual ficam armazenados, às vezes por até muitos anos, até que aconteça de fato a fertilização. Os receptáculos seminais podem ser simples invaginações do exoesqueleto (estruturas externas) ou representados por uma dilatação do oviduto (estruturas internas). Nos decápodes com fertilização interna (braquiúros), os espermatóforos são depositados diretamente dentro de um receptáculo seminal interno. Já nos organismos em que ocorre fertilização externa (maioria dos decápodes), os espermatóforos são colocados na superfície ventral da fêmea ou em um receptáculo externo e a fertilização ocorre à medida que os óvulos vão sendo liberados pelo gonóporos.[3]
Os decápodes aquáticos se atraem por meio de sinais olfativos (feromônios) e tácteis, enquanto espécies terrestres usam sinais tanto visuais quanto auditivos. É comum ocorrer rituais de corte pré-copulatórios. Em muitos decápodes a transferência de espermatóforos do macho para o receptáculo seminal da fêmea só é possível durante um breve momento imediatamente após a fêmea ter sofrido uma ecdise, quando sua cutícula ainda está mole. Em muito braquiúros, o macho carrega a fêmea enquanto espera o momento da muda que se aproxima. Ele a solta enquanto ela muda e a cópula ocorre em seguida.
Sinais visuais e acústicos são de especial importância para a atração nas espécies terrestres. Por exemplo, o caranguejo semiterrestre chama-maré (Uca) exibe um comportamento de corte elaborado que tem sido estudado. Os machos atraem as fêmeas para seus abrigos nos quais deverá ocorrer o acasalamento. As fêmeas permanecem nos abrigos enquanto incubam os ovos até a eclosão das larvas. Os machos desse grupo de caranguejos usam o quelípode maior e muito desenvolvido para atrair as fêmeas e defender seus abrigos contra outros machos. Encontros entre machos dão origem a combates altamente ritualizados, nos quais a quela maior é posicionada como um escudo. Os movimentos de combate envolvem variações de empurrões.[3]
O macho de chama-maré atrai a fêmea movimentado a quela maior em um padrão que é específico para cada espécie. Machos podem também atrair as fêmeas com sinais acústicos que são produzidos com o batimento da quela maior contra o substrato ou flexionando rapidamente as pernas locomotoras. O número de batidas em série e o intervalo entre as séries são específicos para cada espécie. Os sons são transmitidos na forma de vibrações através do sedimento. As fêmeas podem detectar esses sinais a até um metro de distância por órgãos especializados localizados nas pernas.
Durante a cópula, os camarões orientam-se tipicamente em ângulos retos, de tal forma que as respectivas regiões genitais nas superfícies ventrais de cada um fiquem em aposição. O primeiro e o segundo pleópodes do macho encontram-se modificados e são usados na transferência do espermatóforo para um receptáculo localizado entre as pernas torácicas da fêmea.[3]
Em lagostins e lagostas, o macho vira a fêmea de cabeça para baixo e imobiliza os quelípodes dela com os seus próprios. Nos lagostins, os gonópodes são inseridos dentro do receptáculo seminal e os espermatóforos são conduzidos pelos sulcos presentes nos próprios gonópodes. Caso o receptáculo seminal esteja ausente, como acontece nas lagostas, os espermatóforos são presos ao corpo da fêmea, particularmente nas bases dos dois últimos pares de pernas. Esses espermatóforos se apresentam como manchas de cor escura e visíveis nas fêmeas da lagosta do gênero Panulirus. Os anomuros ermitãos precisam sair, em parte, de suas conchas para acasalarem. Os parceiros fazem contato entre suas respectivas superfícies ventrais e os espermatóforos e óvulos são liberados ao mesmo tempo.
Durante a cópula dos caranguejosbraquiúros, um dos indivíduos fica sob o outro, de forma que suas superfícies ventrais fiquem em posições opostas. O abdome do macho, que costuma estar flexionado sob o cefalotórax, é distendido para expor os gonópodes, esses conduzem os espermatóforos são depositados no receptáculo seminal. Em espécies sem receptáculos seminais, os ovos são depositados logo após a cópula. Em espécies com um receptáculo seminal interno e fertilização interna (como os braquiúros), a abertura do receptáculo é selada por um tampão e a fertilização pode não acontecer antes de um ou mais anos após a cópula.
Embora o sexo dos organismos seja determinado geneticamente na maioria dos crustáceos, muitas etapas da reprodução, como a maturação, manutenção e funcionamento das gônadas e o desenvolvimento dos caracteres sexuais secundários, estão sob controle hormonal.[49]
A glândula do seio libera o hormônio inibidor da gônada (GIF) que inibe a maturação dos óvulos durante os períodos não reprodutivos do ano. Durante a estação reprodutiva, o hormônio liberado é o hormônio estimulador da gônada (HEG) a partir de comandos do sistema nervoso central. Na presença desse segundo hormônio, o nível de HIG no sangue diminui e o desenvolvimento dos óvulos começa. Ao mesmo tempo, e também sobre a influência do HEG, o ovário produz um terceiro hormônio que induz alterações anatômicas secundárias necessárias que estão associadas com a incubação dos ovos, como o desenvolvimento de cerdas ovígeras ou o desenvolvimento dos oostegitos nos peracáridos. Essas características surgem com a muda seguinte.
Nos malacóstracos, o desenvolvimento dos testículos e dos outros caracteres sexuais nos machos é controlado por hormônios produzidos em uma massa de tecido endócrino secreto que é chamado de glândula androgênica. Essa glândula se localiza no final do canal deferente ou, até mesmo, nos próprios testículos. Ela é inibida por neurossecreções via glândula do seio, mas é estimulado por neurossecreções do cérebro. Ao ser estimulada, a glândula androgênica secreta o hormônio androgênico, o qual induz à espermatogênese e ao desenvolvimento das estruturas reprodutivas dos machos. Também estimula o comportamento reprodutivo do macho e o desenvolvimento dos caracteres sexuais secundários. A remoção dessa glândula leva à perda das características masculinas e a conversão dos testículos em tecido ovariano. Ao passo que se a glândula for transplantada para uma fêmea, os ovários.[3]
Camarões peneídeos primitivos e táxons relacionados liberam seus ovos diretamente no oceano e os abandonam. Mas, em todos os outros decápodes, os ovos são aderidos e incubados nas fêmeas. Em fêmeas de caranguejos, o abdome, que comumente está flexionado firmemente, é distendido para dar espaço à massa de ovos.
Fase larval Zoéa da espécie Homarus gammarus (Malacostraca, Decapoda, Nephropidae).
O estágio de eclosão varia muito nos decápodes. Nos camarões primitivos, que liberam os ovos direto na água, a forma de eclosão é uma larva náuplio ou metanáuplio. Já em todos os outros decápodes, os ovos são incubados nos pleópodes das fêmeas e, nas espécies marinhas, a eclosão da larva ocorre no estágio de protozoé ou zoé. Os estágios de zoé da maioria dos caranguejos são facilmente reconhecíveis pela presença de um espinho rostral muito longo e algumas vezes por um par de espinhos laterais na margem posterior da carapaça. Ao menos em algumas espécies, os espinhos parecem contribuir para a redução da predação das larvas por pequenos peixes. O estágio de pós-larva dos caranguejos é uma megalopa com todos os pares de apêndices já presentes.[3]
Muitos decápodes com larvas planctônicas habitam águas costeiras rasas, estuários ou manguezais durante a fase adulta ou no estágio de jovem. Por exemplo, os estuários e os manguezais são berçários para muitas espécies de camarões peneídeos. Há uma tendência de abreviação da fase larva nos decápodes que habitam oceanos frios ou profundidades abissais. Nos decápodes estritamente dulciaquícolas, o desenvolvimento é direto e estágios larvais geralmente são ausentes.
O desenvolvimento em quase todos os anomuros e braquiúros terrestres ocorre no mar. Carregando os ovos em desenvolvimento no abdome, a fêmea migra para a costa e entra na água para liberar as zoés recém-eclodidas. O desenvolvimento nesse caso é direto nos caranguejos potamídeos terrestres, os quais são independentes do oceano, mas voltam à água doce para se reproduzir.
Muitos crustáceos, como os lagostins, lagostaHomarus e as cracas, sofrem mudas e crescem durante a vida toda, embora os períodos de intermuda tornem-se progressivamente mais longos.[57] Tais crustáceos podem ter uma vida longa e outros podem se tornar bastante grandes. Em outros, como alguns caranguejos, os processos de muda e crescimento cessam após determinado número de ínstares, quando o indivíduo atinge a maturidade sexual ou um determinado tamanho.
Os processos fisiológicos envolvidos na muda são regulados por interações hormonais e são semelhantes àquelas que ocorrem nos insetos. Um órgão produz ecdisona, que atua nas célulasepidérmicas e cecos digestivos, iniciando a pró-ecdise. A produção de ecdisona é inibida por um hormônio liberado pela glândula do seio, a ação desse hormônio é uma importante diferença com relação ao controle da muda nos insetos, nos quais o corpo cardíaco estimula, ao invés de inibir, a produção de ecdisona.[3]
A regulação dos hormônios da muda, e consequentemente de todo o ciclo de muda, depende de estímulos diferentes que atuam no sistema nervoso central. Nos lagostins, os quais mudam sazonalmente, o fotoperíodo (duração do dia) é o fator de controle,enquanto no caranguejo Carcinus, o crescimento dos tecidos é o fator de controle.
Camarão Peisos petrunkevitchi, da família Sergestidae (Decapoda, Dendrobranchiata)
É composta pelas superfamílias Penaeoidea e Sergestoidea, que possuem camarões como integrantes, e possuem como sinapomorfia exclusiva a presença de dendrobrânquias. Possuem quela nos três primeiros pereópodes, órgãos copuladores modificados no primeiro par de pleópodes nos machos e expansões em direção ventral dos tergitos abdominais. As fêmeas deste grupo não incubam ovos e a fertilização é externa.[58]
Os camarões possuem um tegumento fino e um corpo dividido em duas regiões: cefalotórax e abdômen, com olhos pedunculados e apresentando um par de antênulas, um par de mandíbulas e dois pares de maxilas. O tórax possui três pares de maxilípedes e cinco pares de pereópodes. Os cinco primeiros somitos abdominais apresentam apêndices especializados para a natação e no sexto os apêndices estão modificados como um leque caudal, formado por um par de urópodes e um telson terminal.[59]
Camarão (Penaeus monodon) representante da ordem Decapoda, Dendrobranchiata, Penaeoidea, muito encontrado na Ásia e Austrália
A superfamília Penaeoidea é composta por camarões marinhos, representando aproximadamente um terço das espécies de importância comercial no mundo.[60][61] É um grupo diverso, com mais de 400 espécies distribuídas globalmente e habitando tanto águas rasas quanto zonas abissais abaixo de 5000 metros, ocupando diferentes níveis tróficos da cadeia alimentar em diversas profundidades do oceano.[62]
Camarão do gênero Nematopalaemon representante da ordem Decapoda, Pleocyemata, Caridae
Representantes desse táxon não possuem dendrobrânquias. Os embriões são incubados nos pleópodes das fêmeas e eclodem em estágios tipicamente de zoea.[63][58]
Mais diversos em águas tropicais rasas.[64] Existem cerca de 2500 espécies viventes, são conhecidas pela designação geral de camarões carídeos.[65] Possuem filobrânquias e geralmente o primeiro ou os primeiros pares de pereópodes são quelados e de tamanhos variados. A segunda “pleura” abdominal é marcadamente mais expandida e recobre parcialmente a primeira e a terceira pleura. O primeiro par de pleópodes é geralmente um pouco mais reduzido, mas não muito modificado, nos machos.[58]
Camarão-palhaço representante da ordem Decapoda, Pleocyemata, Strenopodidea
Lagosta da ordem Decapoda, Pleocyemata, Thalassinidea em vista frontal como representante da classe de malacostracos
A maioria das espécies é tropical e associada a ambientes bentônicos, particularmente com recifes de coral, podendo ser comensais, como é o caso de camarões pertencentes ao gênero Stenopus, que podem remover parasitas de peixes.[66] Se caracterizam por possuírem os três primeiros pares de pereópodes são quelados e o terceiro é significativamente maior do que os outros. As brânquias são do tipo tricobrânquia e o primeiro par de pleópodes é unirreme tanto nos machos quanto nas fêmeas, mas não é marcadamente modificado. Apresentam entre 2 a 7 centímetros de comprimento.[58]
Integrantes desse grupo são todos marinhos, como por exemplo as lagostas, apresentando importância econômica[67] possuem como características o abdômen achatado, portador de leque caudal, a carapaça cilíndrica ou deprimida dorso-ventralmente e as brânquias do tipo tricobrânquia. O número de pereópodes quelados é variável, podendo apresentar os quatro primeiros pares, apenas o último ou até mesmo nenhum.[58]
Tem como representantes lagostins e a lagosta norte-americana, podendo ser marinhos ou viver em água doce, mas também em solos úmidos em que é possível a escavação de sistemas complexos e extensos de galeria.[68] O abdômen deprimido dorso-ventralmente termina em um leque caudal bem desenvolvido. Possui brânquias do tipo tricobrânquia e os primeiros três pares de pereópodes são sempre quelados, enquanto que o primeiro par é muito desenvolvido.[58]
O grupo é composto principalmente por camarões escavadores.[69] Apresentam abdômen simétrico e achatado dorso-ventralmente, terminando posteriormente em um leque caudal bem desenvolvido. A carapaça é ligeiramente comprida lateralmente e as brânquias são do tipo tricobrânquia. Os dois primeiros pares de pereópodes são quelados, sendo geralmente o primeiro muito mais desenvolvido.[58]
Os indivíduos do grupo Brachyura são denominados caranguejos verdadeiros. A maioria das espécies é marinha, mas existem espécies dulcícolas, semiterrestres e terrestres de ambientes úmidos.[70] Esses caranguejos apresentam o abdômen simétrico, mas muito reduzido e flexionado sob o tórax, com os urópodes geralmente ausentes. O corpo, recoberto por uma carapaça bastante desenvolvida, é deprimido dorso-ventralmente e geralmente está expandido lateralmente.[71] Filobrânquias estão presentes e o primeiro par de pereópodes é quelado e geralmente muito desenvolvido. Os pereópodes do segundo ao quinto par são tipicamente estenopódios simples e utilizados para andar. Os olhos são posicionados externamente em relação às antenas.[58]
Neste grupo estão os ermitões, porcelanídeos, galateídeos, tatuíras e tatuís. O abdômen pode ser descalcificado e torcido de maneira assimétrica nos ermitões ou simétrico, curto e flexionado sob o tórax nos porcelanídeos. Os ermitões, por possuírem o abdômen torcido, podem ocupar conchas de gastrópodes.[72] A carapaça e estruturas branquiais variam, porém o primeiro par de pereópodes é quelado e o terceiro par nunca é quelado; o segundo, quarto e quinto par geralmente são simples, mas podem ser quelados ou subquelados em alguns casos. Geralmente o quinto par de pereópodes é muito reduzido e não é utilizado como apêndices locomotores, mas sim como responsável pela limpeza das brânquias. Os olhos estão posicionados na região interna ao segundo par de antenas.[58]
Estratégias adaptativas associadas à colonização do ambiente de água doce
Com base em análises filogenéticas, a paleontologia nos revela que os decápodes podem ter se originado há cerca de quatrocentos milhões de anos. A irradiação desse grupo teve início no período Mesozoico, que corresponde a aproximadamente 251 a 65,5 milhões de anos atrás.
Menos de 20% de todas as espécies viventes de Decapoda conquistaram com sucesso ambientes salobros e de água doce.[73] O padrão de distribuição dos decápodes lacustres é derivado de inúmeras invasões ao ambiente, favorecidas ao longo dos anos pela seleção natural, como é evidenciado no registro fóssil e em dados moleculares. Evolutivamente, as espécies de água doce descendem de espécies marinhas e tal adaptação pode ser mensurada não apenas considerando os mecanismos osmorregulatórios, mas também os aspectos reprodutivos e do fenômeno de socialização, como o número e tamanho dos ovos, desenvolvimento larval (com abreviação ou supressão) e cuidado parental.[73][74]
Muitas espécies de decápodes de água doce pertencem à família Aeglidae, que compreende 3 gêneros, Aegla, Protaegla e Haumuriaegla. Desta família, apenas o gênero Aegla é vivente atualmente, e os outros dois gêneros foram extintos e são conhecidos apenas através de fósseis de sedimentos marinhos, o que indica a origem marinha do grupo.
O gênero Aegla é constituído por cerca de 69 espécies, todas adaptadas à vida em água doce e endêmicas de áreas temperadas e subtropicais da América do Sul. Elas foram agrupadas filogeneticamente em cinco grandes clados, refletindo, em grande parte, sua distribuição geográfica no sul da América do Sul.[75][76]
A colonização do ambiente de água doce pelos Decapoda exigiu diversas adaptações fisiológicas, reprodutivas, comportamentais, etc. Com isso, observamos nestes animais algumas características únicas que os diferenciam de outros decápodes.
A maioria dos grupos de decápodes marinhos são osmoconformadores, regulando a concentração do seu sangue dentro de uma faixa estreita em torno da concentração de água do mar (~1000 mOsm/kg). Variações na salinidade podem causar alterações fisiologicamente perigosas na concentração iônica dos fluidos corporais extracelulares (a hemolinfa), na osmolaridade do citosol e no volume celular de decápodes aquáticos.
As concentrações de íons orgânicos na água doce são muito menores do que no ambiente marinho. Isso representa uma mudança drástica nas condições ambientais, uma vez que a disponibilidade de minerais essenciais e a pressão osmótica são reduzidas. Essa mudança demanda tanto adaptações fisiológicas quanto comportamentais.
Habilidades osmorreguladoras como: a presença de uma cutícula espessa com permeabilidade reduzida a íons e água, estruturas celulares e teciduais altamente especializadas e bombas metabólicas ativas que movem íons no corpo são adaptações evolutivas seletivamente vantajosas observadas em decápodes juvenis e adultos de água doce. Tais características permitem que os decápodos que vivem em água salobra e doce se oponham à diluição de seus fluidos corporais do ganho de água e da perda de íons.
Os processos de osmorregulação intracelular causam perdas metabólicas significativas de matéria orgânica e energia e podem compensar apenas pequenas flutuações na salinidade do habitat. A regulação iônica extracelular é um mecanismo energeticamente mais eficiente, que consiste em um conjunto complexo de processos ativos que promovem o transporte de íons através das membranas celulares em células epidérmicas altamente especializadas chamadas ionócitos. A regulação iônica extracelular é tipicamente encontrada em habitats de água doce, estuarinos e terrestres.
Quando os decápodes vivem em meio diluído (água salobra ou doce), eles osmorregulam de modo que as concentrações de íons no espaço da hemolinfa extracelular são reguladas positivamente para compensar as perdas passivas de íons osmóticos e a entrada de água.
Decápodes do gênero Aegla possuem respiração tricobranquial, o que os separa das famílias de Galatheoidea que possuem filobrânquias, vistas em diversos anomuros marinhos. As tricobrânquias dos aeglídeos de água doce possuem uma estrutura única. Os filamentos das brânquias são longos tubos semelhantes a dedos, estes se estendem para frente a partir da base da brânquia. A porção proximal da brânquia assemelha-se à de uma filobrânquia de braquiúro por ter lamelas em forma de placa que se estendem sobre o eixo branquial.[77]
Desenvolvimento pós-embrionário, cuidado parental e ciclo de vida
O desenvolvimento padrão ontogenético dos crustáceos compreende três fases: (1) desenvolvimento dentro de um ovo, (2) eclosão da larva na forma de náuplio seguido por larvas planctônicas de vida livre, e (3) transformação da larva em um juvenil, que crescerá para a reprodução.
Uma característica singular para os crustáceos é a fase larval, pois a mesma reflete a ontogenia do grupo e pode conter traços específicos para algumas espécies, de diferentes famílias, sendo assim alvo de estudos.
A abreviação ou supressão do estágio larval (desenvolvimento direto) e a extensão do cuidado parental são características importantes para o sucesso evolutivo dos decápodes de água doce. Por exemplo, espécies do gênero Macrobrachium, que vivem em águas salobras, estuários, lagoas costeiras e cabeceiras de rios, têm mostrado uma impressionante relação entre adaptação lacustre e abreviação do estágio larval. O maior exemplo encontrado neste gênero é a espécie Macrobrachium dayanum, que possui um estágio larval bentônico de vida livre protegido por sua mãe nos primeiros dias após a desova, o que contrasta muito bem com o padrão de estágios larvais encontrado na família, que concentra-se entre nove e onze estágios zoécios.[73]
Para haver um sucesso evolutivo no desenvolvimento direto, é necessário um sistema de fixação de ovos altamente eficaz. Em Peracarida, por exemplo, tal desenvolvimento foi propiciado graças a “berços” no corpo da fêmea conhecido como “câmara de incubação”; nos caranguejos de água doce, nos lagostins e nos caranguejos primários de água doce, o cuidado parental é obrigatório e realizado sob o pleón (abdômen); nos caranguejos secundários de água doce, o cuidado parental é menos frequente, mas bastante variável, incluindo o transporte das crias nos pleópodes ou em cima da carapaça e a criação de larvas e juvenis livres em habitações parecidas com ninhos; nos aeglídeos, os ovos são carregados nos quatro pares de pleópodes e são incubados por um período de quatro a oito meses.[78]
O desenvolvimento pós-embrionário dos aeglídeos de água doce (gênero Aegla), por exemplo, diferentemente dos anomuros marinhos, é direto em 75% dos casos, o que significa a supressão total da fase larval e eclosão de juvenis bentônicos que, de maneira geral, se assemelham morfologicamente aos indivíduos adultos, embora os traços morfológicos associados aos estágios zoea e megalopa ainda possam ser reconhecidos dentro do ovo em desenvolvimento.[73][79][80][81][82] Isso é conseguido, como dito anteriormente, transportando os embriões nos pleópodos maternos até a eclosão deles como juvenis, ou pela incubação de estágios zoea nascidos antes, ou nos pleópodos maternos, ou em poças ou tocas de viveiros de água doce até que o estágio juvenil seja atingido.[73] Esses dados contrastam muito com os decápodes marinhos, nos quais este tipo de desenvolvimento só foi descrito em cerca de 20 a 30 espécies.[73][80]
Cerca de 15 a 20% dos decápodes de água doce apresentam desenvolvimento indireto com estágio larval abreviado.[73] Outros decápodes que apresentam desenvolvimento indireto possuem estágio larval longo que necessita de água salgada, pois as larvas ainda não possuem as adaptações fisiológicas necessárias para sobreviver às condições de estresse osmótico.[73][82]
O desenvolvimento indireto com fase larval prolongada implica, em algumas espécies, a necessidade das larvas serem transportadas para o mar ou para águas estuarinas ou costeiras para se desenvolverem. Este transporte da água doce para a salgada, chamado de migração diádroma, também faz parte de uma estratégia adaptativa que permite a vida adulta em habitats não marinhos, em combinação com a conservação do padrão ancestral da história de vida (desenvolvimento larval planctônico prolongado em água salgada). Os estágios larvais tardios ou pós-larvais precoces voltam para a água doce por conta própria, onde se desenvolvem em juvenis e adultos.[82]
Espécies que vivem parte do seu ciclo de vida em água salgada e parte em água doce, mas não especificamente por razões reprodutivas (como no caso das migrações diádromas), e sim fisiológicas, são chamadas de anfídromas, como é o caso das larvas que não possuem capacidades osmorreguladoras para viver em água doce e vivem em água salgada até atingirem a maturidade. Neste caso, os indivíduos crescem, se reproduzem e depositam os ovos em lagos ou rios, mas as larvas se desenvolvem em estuários marinhos ou em áreas marinhas costeiras. Ao completar o desenvolvimento larval, os indivíduos devem migrar rio acima até o habitat de água doce, onde viverão toda a vida adulta. Um benefício importante da anfidromia é a dispersão das larvas marinhas através dos rios. Por isso, as espécies anfidrômicas apresentam uma distribuição geográfica mais ampla do que as espécies de água doce sem larvas marinhas.
Além das diferenças de desenvolvimento, existem diferenças marcantes no número e tipo de ovos produzidos entre decápodes marinhos, de água doce e espécies anfídromas.[73] Decápodes marinhos produzem um número de ovos que vai de milhares até milhões, dependendo da espécie. As espécies de decápodes de água doce produzem um número de ovos muito menor, que vai de dezenas a centenas, e poucas vezes chega aos milhares, e, neste caso, os ovos são geralmente maiores e com mais vitelo, para sustentar a intensa incubação e o desenvolvimento do embrião antes dos ovos eclodirem, e os indivíduos costumam eclodir como juvenis já parecidos com os adultos ou como larvas em estágio avançado que terão poucos estágios subsequentes. Já no caso de espécies anfídromas, as fêmeas botam muitos ovos pequenos, que eclodem em um estágio larval muito menos avançado em relação às espécies exclusivamente de água doce (análises comparativas podem ser feitas observando a Tabela 1).
A diminuição na quantidade e no tamanho dos ovos é uma consequência explicada pela “hipótese do porto seguro”, que diz que a seleção natural tende a maximizar a quantidade de tempo que os organismos passam em estágios de desenvolvimento mais seguro e minimizando estágios onde há maior taxa de mortalidade pode ser mais alta. Logo, tais eventos (abreviação do estágio larval e extensão do cuidado parental) compreendem um “trade-off”, pois há um maior investimento energético, em contrapartida, há uma diminuição da predação. Ou seja, o gasto é compensado por uma alta fecundidade e futura reprodução, subsidiando influências filogenéticas e biogeográficas, agindo no padrão de dispersão das espécies.
Algumas espécies de decápodes de água doce não fazem migrações diádromas, mas liberam suas larvas no habitat adulto límnico.[83] Nestas espécies, as necessidades das larvas por sais e alimento planctônico requerem tempos de transporte rápidos para atingir águas estuarinas ou costeiras. Assim, o habitat adulto deve estar localizado em rios de fluxo rápido e não em águas interiores estagnadas, e a distância entre o habitat adulto e o mar deve ser muito mais curto do que em espécies com migrações adultas.[82]
As espécies que não fazem migrações diádromas e possuem um ciclo de vida bifásico não são completamente adaptadas aos seus habitats, podendo ser evolutivamente transitórias ou instáveis, e tendendo a desaparecer com o tempo. Porém, perdas de traços reprodutivos e de desenvolvimento previamente vantajosos são provavelmente compensadas por ganhos evolutivos de outras características, como o desenvolvimento precoce dos órgãos osmorregulatórios, a independência parcial ou completa das larvas de fontes alimentares planctônicas e o cuidado materno que reduz a mortalidade larval.[82]
Além disso, algumas espécies de decápodes de água doce possuem estágios larvais que podem sobreviver e se desenvolver em água doce e não precisam ser exportadas para ambientes de água salgada. Isto requer adaptações, em particular capacidades osmorreguladoras, que permitam a sobrevivência e desenvolvimento em água doce ao longo do ciclo de vida. A grande maioria desses decápodes tem uma fase larval curta, que é intermediária entre um ciclo de vida bifásico com um desenvolvimento planctotrófico estendido e um ciclo monofásico sem larvas.[82] Tais espécies mostram uma tendência à lecitotrofia, ou seja, não procuram o alimento no meio externo, mas se alimentam a partir das reservas de vitelo que levam consigo desde seu nascimento. A alta concentração de carbono e a alta relação C:N nos ovos indica que o estoque de lipídio no vitelo é proporcionalmente maior que a quantidade de proteína, o que provide um substrato bioquímico energético para a independência de alimento em relação ao meio externo. Além disso, tais espécies apresentam estágio larval zoea que pode continuar seu desenvolvimento até a próxima fase na ausência completa de comida.[84]
A lecitotrofia, citada anteriormente, apresenta algumas desvantagens. Por se basear no aumento do investimento em energia materna por filhos, implica a redução ou na não exploração de fontes alimentares planctônicas. Com isso, as larvas eclodem com grande tamanho corporal e alto teor de gordura, e mostram tipicamente pouca ou nenhuma atividade de natação, tornando-se atraentes para predadores e reduzindo seu potencial de dispersão, a colonização de novos habitats ou a substituição de populações anteriormente extintas.[82]
Além disso, o cuidado parental é um padrão frequentemente observado em decápodes de água doce, ocorrendo em cerca de 70% das espécies.[73] Este cuidado se estende até a fase juvenil bentônica, e com isso os primeiros estágios pós-embrionários permanecem protegidos do estresse físico ou nutricional no ambiente pelágico.[82] Nos Aeglidae, os ovos são carregados nos 4 pares de pleópodos maternos, incubados por cerca de 4 a 8 meses, e os juvenis, que já possuem morfologia parecida com os adultos, permanecem agarrados nos pleópodos maternos por mais alguns dias após a eclosão.[73]
O encurtamento ou supressão total do estágio larval, a extensão do cuidado parental pós nascimento e a dispersão juvenil limitada são importantes estratégias observadas em diversos decápodes e outros animais de água doce, como peixes e gastrópodes, e essas características estão relacionadas com o sucesso da colonização desses habitats.[73][82] Além disso, essas características estão associadas à dispersão e fluxo gênico reduzido entre as populações destas espécies, o que pode explicar o alto grau de endemismo e especiação desses decápodes.[82]
Outro fator importante no ciclo de vida dos aeglídeos de água doce é que o período reprodutivo dos mesmos ocorre na estação fria e seca do ano, quando as velocidades e fluxo da chuva são menores. Isso garante que os juvenis serão grandes o suficiente para suportar as correntes de água mais fortes no pico da estação chuvosa e quente que ocorre 6 meses depois.[76][85][86]
Tabela 1. Exemplos de cuidado parental pós desova e prolongamento da proteção parental em Decápodas lacustres (retirado de forma literal do artigo do Vogt, 2013[73])
Espécies
Família
Duração
Referência
Caridea
Dugastella marocana (Bouvier)
Atyidae
n.d
Huguet et al. (2011)
Dugastella valentina (Ferrer Galdiano)
Atyidae
5–9 d
Cuesta et al. (2006b)
Astacida
Astacus astacus (Linnaeus)
Astacidae
6–17 d (a)
Baumann (1932) and Bieber (1940)
Astacus leptodactylus (Eschscholtz)
Astacidae
20–25 d
K¨oksal (1988)
Austropotamobius pallipes (Lereboullet)
Astacidae
25 d
Gherardi (2002)
Pacifastacus leniusculus leniusculus (Dana)
Astacidae
∼20 d
Andrews (1907)
Pacifastacus leniusculus trowbridgii (Stimpson)
Astacidae
15–19 d
Mason (1970b)
Cambaroides japonicus (De Haan)
Cambaridae
n.d.
Scholtz & Kawai (2002)
Cambaroides similis (Koelbel)
Cambaridae
6 sem. (a)
Ko & Kawai (2001)
Orconectes neglectus chaenodactylus (Williams)
Cambaridae
20 d
Price & Payne (1984)
Orconectes limosus (Rafinesque)
Cambaridae
16–24 d
Andrews (1904) and Mathews (2011)
Orconectes pellucidus (Tellkampf)
Cambaridae
27–40 d
Bechler (1981)
Procambarus fallax (Hagen) f . virginalis
Cambaridae
14–38 d
Vogt (2008)
Procambarus clarkii (Girard)
Cambaridae
∼3 sem. a 3 meses (b)
Huner (1988) and Gherardi (2002)
Cherax cainii (Austin & Ryan)
Parastacidae
28–38 d
Burton et al. (2007)
Cherax destructor (Clark)
Parastacidae
21–22 d (a)
Sandeman & Sandeman (1991)
Cherax quadricarinatus (Von Martens)
Parastacidae
18–29 d
Levi et al. (1999)
Engaeus cisternarius (Suter)
Parastacidae
73–95 d
Suter (1977)
Engaeus orramakunna (Horwitz)
Parastacidae
>1 ano (b)
Duffy (2010)
Ombrastacoides pulcher (Riek)
Parastacidae
1.5–2 meses, 12 meses (b)
Hamr & Richardson (1994)
Astacopsis gouldi (Clark)
Parastacidae
44–58 d (a)
Hamr (1992)
Paranephrops planifrons (White)
Parastacidae
40–50 d
Hopkins (1967)
Paranephrops zealandicus (White)
Parastacidae
4–5 meses
Whitmore & Huryn (1999)
Virilastacus araucanius (Faxon)
Parastacidae
20 d
Rudolph & Rojas (2003)
Anomura
Aegla castro (Schmitt)
Aeglidae
∼15 d
Swiech-Ayoub & Masunari (2001)
Aegla platensis (Schmitt)
Aeglidae
5 d
L´opez Greco et al. (2004)
Aegla perobae (Hebling & Rodrigues)
Aeglidae
8–12 d
Rodrigues & Hebling (1978)
Aegla uruguayana (Schmitt)
Aeglidae
3–4 d
L´opez Greco et al. (2004)
Brachyura
Candidiopotamon rathbunae (De Man)
Potamidae
9–16 d
Liu & Li (2000)
Potamon fluviatile (Herbst)
Potamidae
n.d.
Pace et al. (1976)
Sinopotamon yangtsekiense (Bott)
Potamidae
n.d.
Xue et al. (2010)
Liberonautes latidactylus (De Man)
Potamonautidae
n.d.
Cumberlidge (1999)
Platythelphusa praelongata (Marijnissen et al.)
Potamonautidae
n.d.
Marijnissen et al. (2004)
Potamonautes lirrangensis (Rathbun)
Potamonautidae
algumas semanas
Rademacher & Mengedoht (2011)
Sudanonautes africanus (Milne-Edwards)
Potamonautidae
n.d.
Disney (1971)
Kingsleya ytupora (Magalhães)
Pseudothelphusidae
n.d.
Wehrtmann et al. (2010)
Austrothelphusa transversa (Von Martens)
Gecarcinucidae
alguns dias
Powers & Bliss (1983)
Spiralothelphusa hydrodroma (Herbst)
Gecarcinucidae
38–100 d (b)
Pillai & Subramoniam (1984)
Parathelphusa maculata (De Man)
Gecarcinucidae
n.d.
Rademacher & Mengedoht (2011)
Sylviocarcinus pictus (Milne Edwards)
Trichodactylidae
n.d.
Souza da Silva (2010)
Geosesarma peraccae (Nobili)
Sesarmidae
8–9 d (c)
Lam (1969)
Geosesarma notophorum (Ng & Tan )
Sesarmidae
4 d
Rademacher & Mengedoht (2011)
Geosesarma bicolor (Ng & Davie)
Sesarmidae
alguns dias
Rademacher & Mengedoht (2011)
Metopaulias depressus (Rathbun)
Sesarmidae
3 meses (c), >1 ano (d)
Diesel & Schubart (2007)
Sesarma jarvisi (Rathbun)
Sesarmidae
2–3 meses (c)
Diesel & Horst (1995)
*a coluna de duração denota ao tempo de carregamento em estágios de pós desova em pleópodes maternais (período de anexamento somado de período de descarregamento), se não indicado de outra forma. “N.d.”, significa não determinado, d. dia(s).
*a. Apenas estágio de anexação
*b.Transporte de jovens e posterior proteção de juvenis livres em toca
*c. Brotamento da larva livre e estágios pós larvais em cuidados enterrados ou (water-filled:gente como traduzo isso de forma que dê sentido?)
*d. Proteção de estágios pós larvais livres em grupos de famílias eusociais
A mudança evolutiva entre um desenvolvimento larval longo para o desenvolvimento direto pode ter resultado em consequência consideráveis para a biogeografia do táxon, porque espécies de desenvolvimento direto possuem capacidades de dispersão reduzida quando comparados a espécies de desenvolvimento planctônico.[73] A maioria das espécies de decápodes de água doce de desenvolvimento direto têm uma faixa geográfica estreita, enquanto este não é o caso em espécies anfídromas.[87][88][89][90][91]
A baixa capacidade de dispersão é um importante fator na evolução do endemismo, logo, espécies endêmicas são comuns nos casos de decápodes de desenvolvimento direto.[73]
Segundo Hansen (1992),[92] espécies de desenvolvimento direto sofrerão especiação mais rapidamente que espécies que possuem estágio larval planctônico. Isso se deve ao fato de que as distribuições geográficas no primeiro caso são mais restritas, levando ao isolamento reprodutivo e diminuição do fluxo gênico.
Mecanismos de absorção de sal pelo epitélio branquial compensando a perda difusiva para o meio externo
Produção de urina hiposmótica ou isosmótica com reduzido fluxo pelas glândulas antenais
Regulação do volume intracelular através de efetores osmóticos orgânicos pelas espécies recentes invasoras que migram para a água salobra para completar seu ciclo de vida.
Os comportamentos dos pais, em especial das fêmeas, pode ser alterado em detrimento do cuidado da prole. Nos lagostins, tomados como exemplo, além da fixação nos pleópodos (que requer uma série de mudanças rítmicas do lado esquerdo para o direito, pelos ganchos pereopodais), pode ser visto a busca pela territorialidade, defesa da progenitura, ventilação e defesa dos ovos. Para os machos, os estudos sobre seus cuidados com a prole ainda carecem de estudo. Mas, ao que se sabe atualmente, os tipos de cuidados dos mesmos são secundários, como tarefas de enterrar e limpar os ovos.
Estado de conservação dos crustáceos decápodes da fauna brasileira
Muitos fatores contribuem para ameaçar o grupo de decápodes ao longo de sua distribuição, como declínio da qualidade de seu habitat, contaminação da água pelo uso de pesticidas ou por esgoto humano, silvicultura, construção de usinas elétricas, entre outros.[93] A ordem Decapoda é a mais diversificada dentre o subfilo Crustacea (classe Malacostraca) apresentando espécies de grande interesse econômico como camarões, lagostas e caranguejos.[94]
Dentre os grupos que se encontram com considerações em listas vermelhas Malacostraca possui 3016 espécies listadas (em 2019), sendo que Decápodes contabilizam mais de 95% de representatividade nos índices de conservação dessa classe, apresentando 2892 espécies listadas. Além disso, 1130 espécies da ordem mencionada estão sob o rótulo de Dados Insuficientes, ou seja, a situação de conservação de boa parte de Decápodes encontra-se ainda sob difícil determinação.[95]
Relatos históricos e registros iconográficos da fauna de decápodes do Brasil: do descobrimento ao Império
Reprodução da carta de Pero Vaz de Caminha, a qual descrevia as terras brasileiras durante a expedição de Cabral. Essa reprodução pode ser encontrada na Biblioteca Nacional de Portugal
Pero Vaz de Caminha foi o escrivão da frota portuguesa sob o comando de Pedro Álvares Cabral à chegada em Porto Seguro em abril de 1500. Caminha enviou uma carta a Dom Manuel I — então rei de Portugal — descrevendo diversos aspectos das terras recém-descobertas. Nessa carta, Caminha disserta a respeito da expedição, sobre as características físicas e hábitos culturais dos indígenas; fornecendo, ademais, relatos vagos da fauna e flora observada — o que dificulta tentativas de se identificar as espécies encontradas.[96]
A única referência a crustáceos encontrada na carta de Caminha foi a descrição de camarões do local:
"Enquanto lá estávamos foram alguns buscar marisco e não no acharam. Mas acharam alguns camarões grossos e curtos, entre os quais vinha um muito grande e muito grosso; que em nenhum tempo o vi tamanho."[97]
Para Leitão (1941), há a possibilidade de que os camarões encontrados fossem os de água doce Macrobrachium acanthurus (Wiegmann, 1836) ou os marinhos Squilla (Fabricius, 1787) ou Lysiosquilla sp. (Dana, 1852); isso porque, de acordo com a carta de Caminha, o desembarque ocorrera em área próxima a um rio, não havendo, entretanto, especificações quanto à região de pesca desses animais — se no curso do rio ou na praia[98].
Gabriel Soares de Sousa esteve na costa da Bahia 69 anos depois e, como Caminha, descreveu os camarões do local como "grossos" ou "robustos". Esses camarões eram chamados pelos índios de "potiguaçu" (literalmente, camarão grande e grosso, em tupi[99]). Ilustrações de potiguaçu sugerem que esse camarão possa ser Macrobrachium carcinus (Linnaeus, 1758).[96]
O camarão M. carcinus apresenta a ponta do rostro curvada para cima e possui entre 11 e 14 dentes na margem superior e 3 ou 4 dentes na margem inferior do rostro.[100] O potiguaçu representado em algumas ilustrações apresenta rostro curvado para cima, 11 dentes na margem superior e 4 na margem inferior. Segundo Holthuis (1952), as características que melhor definem o gênero Macrobrachium são o formato do rostro e do segundo par de pernas.[100]
Não há clareza, entretanto, sobre as características do segundo par de pernas nas ilustrações. Porém, o padrão de cores ilustrado no potiguaçu é semelhante ao de Macrobrachium carcinus. A área em que foi encontrado o potiguaçu também coincide com a que é encontrado o camarão M. carcinus, situada entre a Flórida e o sul do Brasil.[96]
José de Anchieta foi um frade jesuíta, nascido em 19 de março de 1534, em São Cristóvão de Laguna, nas Ilhas Canárias. Chegou ao Brasil em 15 de julho de 1553, e em 1560, escreveu uma carta que descreve brevemente parte a natureza de São Vicente (atualmente São Paulo), incluindo mamíferos, pássaros, répteis, peixes, crustáceos, insetos e plantas. Quatro espécies de crustáceo foram descritas: Portunidae sp., Uca sp., U. cordatus (Linnaeus 1763) e Cardiosoma guanhumi (Latreille, 1828). Anchieta organizou a sistematização do tupi, uma síntese de vários dialetos e línguas falados por diversos grupos nativos, que facilitou a comunicação entre os indígenas e os colonizadores. Isso permitiu a redução da discrepância nos registros, feitos por diferentes autores, sobre espécies locais conhecidas pelos nativos.[96]
Os franceses buscaram se estabelecer no Brasil, e Jean de Léry foi um dos homens presentes na primeira expedição. Suas narrativas foram um dos primeiros documentos que descreviam o Brasil. André Thevet, que também estava presente, coletou informações sobre etnologia mas nada foi informado sobre crustáceos em seu trabalho publicado.[96]
Jean de Léry (1534 - 1611) nasceu em La Margelle (Borgonha, França). Aos 18 anos foi para Genebra e se tornou aluno de teologia de João Calvino. Posteriormente, Calvino acabou por receber uma solicitação do almirante Villegagnon para ajudar na empreitada da criação da França Antártica, havendo a possibilidade de propagar sua doutrina pelo Novo Mundo.[101] Enviou uma equipe de 14 pessoas, incluindo Jean de Léry, que seria então o futuro narrador da obra "Viagem à Terra do Brasil", publicada pela primeira vez em 1578.[102] Esse livro descreve aspectos da malograda expedição, incluindo aspectos culturais dos indígenas e algumas poucas informações acerca da fauna e flora local.[96]
Jean de Léry chegou ao Rio de Janeiro em 26 de fevereiro de 1557, retornando a Genebra após 11 meses no Brasil. Inicialmente não publicou notas sobre a viagem, porém foi incentivado por um colega a enviar um manuscrito para publicação. O manuscrito foi perdido e, baseado em seus rascunhos, redige um segundo, que não foi publicado por conta de um contexto de conflitos religiosos entre protestantes e católicos. Em 1578, a primeira versão da narrativa foi, finalmente, publicada na França.[102]
Em seu relato, há apenas uma menção a um crustáceo (decápode), tendo sido relatado um caranguejo chamado pelos indígenas de uçá (ussa), que corresponderia a um Ucides cordatus.[96] Esse caranguejo foi encontrado em uma área de manguezal, tendo sido confirmado posteriormente[103] que a espécie habita a região indicada por Léry. A respeito dessa menção, pode-se ler na narrativa:
"Existem ainda caranguejos terrestres a que os tupinambás chamam ussa, e surgem aos bandos nas praias e outros lugares pantanosos. Quando alguém se aproxima, fogem de costas e se salvam com celeridade nos buracos abertos nos troncos e raízes das árvores, donde não podem ser tirados sem perigo por causa de seus ferrões, embora possa a pessoa chegar facilmente até o buraco visível. Mais magros do que os caranguejos marinhos, quase não têm carne e exalam cheiro de raiz de cânhamo, não sendo de bom paladar".[104]
Gabriel Soares de Sousa nasceu em torno de 1540, possivelmente em Lisboa. Visitou o Brasil durante a expedição de Francisco Barreto para Moçambique. Alguns que estavam na expedição decidiram ficar no Brasil, entre eles Gabriel de Sousa, que viveu na Bahia por 17 anos. Seu irmão João de Sousa viajou por 3 anos pelo interior do Brasil, encontrando ouro, pedras preciosas e fazendo mapas de suas descobertas. Quando João morreu, Gabriel tornou-se dono de seus mapas e isso fez com ele se interessasse pelo estudo da história natural do Brasil.
Sousa foi à Espanha e pediu a Filipe II o apoio para uma expedição para o interior do Brasil, escrevendo, nesse período, o Tratado descritivo do Brasil (1587). Esse trabalho tinha a intenção de mostrar a riqueza presente no Brasil, a importância de expedições para explorar a área e os riscos de invasões estrangeiras. Alguns anos depois, tendo apoio de Filipe II, Sousa chega ao Brasil. Seu livro é um resultado de 17 anos em território brasileiro e trata sobre temas como geografia, mineralogia, zoologia e botânica. Os crustáceos mencionados por Sousa são[96]:
Siris, caranguejos do gênero Callinectes capazes de nadar e que apresentam patas traseiras em forma de remo. Foram encontradas 7 espécies, sendo a mais comum C. danae;
Caranguejos comestíveis chamados de "guaiá" (Menippe nodifrons) e caranguejos utilizados como isca para pesca, chamados de "guaiauça" (Ocypoda quadrata);
Pequenos caranguejos possivelmente pertencentes ao grupo Pinnatheridae;
Camarões de água doce pertencentes ao gênero Macrobrachium com nomes populares de: potiuaçu, poti e araturé;
Assim como a primeira, a segunda tentativa de ocupação fracassou. Porém, trabalhos importantes foram publicados, sendo os mais relevantes em relação a crustáceos publicados por Claude d'Abbeville.
Caranguejo Cardiosoma guanhumi, representado em gravura por Albert Eckhout; encontrado na obra Theatrum Rerum Naturalium Brasiliae.
Claude d'Abbeville embarcou a caminho do Brasil e chegou em Fernando de Noronha. Ficou no local por 15 dias. 5 meses após partir da França, a frota francesa chegou à ilha de Santa Ana, na costa do Maranhão. Claude d'Abbevile esteve no Brasil por quatro meses e sua primeira narrativa foi publicada em 1614, passando por sete reedições, em francês e português.[96]
Seu trabalho conta sobre suas observações dos índios e apresenta temas como botânica, zoologia e geografia. Dedicou quatro capítulos sobre a fauna do local: um sobre aves, um sobre peixes, um sobre "animais terrestres" (mamíferos, répteis e anfíbios) e o último sobre "os animais imperfeitos do Maranhão" (mosquitos, pulgas, formigas).[96]
Sete espécies de crustáceos foram mencionadas, sendo todas de caranguejos (Brachyura). Como não tinha familiaridade com o tupi-guarani, sua grafia dos nomes nativos era bem peculiar. As espécies mencionadas são[96]:
Um caranguejo marinho, azul e amarelo, chamado de "aratou" (aratu). Esse padrão de cores é encontrado em Eriphia gonagra (Fabricius, 1781). Claude d'Abbevile aplicou o nome nativo "aratu" a essa espécie, apesar de ser utilizado em Aratus pisonii (H. Milne Edwards);
Callinectes sp., chamado de "siry" (siri), um caranguejo marinho com as cores azul e branco;
"Aouära-oussa" (grauçá), descrito como um caranguejo branco que cava suas tocas na praia e leva fragmentos de rochas e conchas. É possivelmente um "caranguejo-fantasma", Ocypode quadrata (Fabricius);
As últimas referências foram do caranguejo "ouraroup", que não apresentava um nome representado no tupi-guarani. Era um caranguejo de água doce (possivelmente um Trichodactylidae), maior do que uma mão fechada e predado pelo "oussapeve" (ou uçá-peva, que significa "caranguejo achatado). O caranguejo achatado é, provavelmente, Callinectes sapidus Rathbun, 1895. Esse caranguejo vive em água doce, longe do mar.[96]
Cristóvão de Lisboa nasceu em 25 de julho de 1583, em Lisboa. Ele veio ao Brasil em 1624, chegando ao estado de Pernambuco no dia 2 de maio.
Entre 1624 e 1627, ele escreveu um manuscrito sobre a sociedade e a natureza do Maranhão. Esse manuscrito foi perdido após o terremoto de Lisboa em 1755; o volume que detalha a fauna brasileira foi reencontrado em 1933, porém partes do registro ainda estão faltando. Entre os animais registrados por ele, encontram-se os seguintes decápodes[96]:
Em 1636, o território neerlandês no Brasil foi repassado para a administração do então conde João Maurício de Nassau-Siegen, ocupando os cargos de governador, capitão e almirante general em nome da Companhia das Índias Ocidentais.[107] No ano seguinte (1637), Nassau foi a Pernambuco levando consigo uma comitiva de artistas e naturalistas, para além de soldados e oficiais.[107][106] Governou de 1637 a 1644, exortando nesse período a realização de estudos em campos diversos, tais como: zoologia, botânica, astronomia e etnologia.[106][107] Nesse sentido, o seu séquito de artistas e naturalistas contribuiu extensivamente para a formação de uma história natural brasileira, pois apresentavam interesses diferentes dos exploradores pretéritos, produzindo documentos detalhados sobre os elementos naturais locais.[96] Dentre eles, destaca-se o papel dos naturalistas Guilherme Piso e George Marcgraf e os artistas Albert Eckhout e Frans Post.[106]
A partir de 1640, com o fim da União Ibérica, a ocupação holandesa começou a enfraquecer política e economicamente. Tornando-se independente da Espanha, Portugal voltou-se para os seus domínios no Brasil.[106] Em 1644, Nassau não obteve apoio militar e financeiro da Companhia das Índias Ocidentais e acabou retornando para a Holanda. Portugal retorna, posteriormente, seu controle sobre Pernambuco e a Companhia acabou por sair do Brasil em 1661.[96]
Laet contribuiu também para a realização e divulgação de outros estudos sobre o Brasil, tendo sido responsável pela organização e publicação da Historia Naturalis Brasiliae (História Natural do Brasil), a partir de manuscritos de Piso e Marcgraf.[107]
Theatrum Rerum Naturalium Brasiliae (Teatro das Coisas Naturais do Brasil)
Gravura de camarões potiatinga e potiguaçu, de autoria atribuída a Albert Eckhout, retirado do Theatrum Rerum Naturalium Brasiliae. O camarão potiguaçu pode ser representado pela coloração amarronzada.
No século XIX, por volta de 1828, as aquarelas e pinturas a óleo produzidas pelos artistas da missão de Nassau entre 1660 e 1664 (no esforço de documentar o que encontraram no Novo Mundo) foram compiladas com outra coleção de ilustrações, denominada Libri Picturati — o conjunto perfazendo 144 volumes.[96][106] As imagens referentes ao Brasil, denotando cerca de duas mil representações diversas de plantas e animais. passaram a ser referenciadas como Libri Picturati A 32-38, na qual se encontra o Theatrum Rerum Naturalium Brasilae (Teatro das Coisas Naturais do Brasil[107]). Esse é subdividido em quatro volumes (A 32-35),[96][106] a saber:
Icones Aquatilium (A 32):
Apresentam 44 ilustrações de peixes e 25 de crustáceos, no que se incluem 13 espécies diferentes de decápodes, uma de stomatopoda e duas de cracas;
Apresenta 172 ilustrações de plantas, flores e frutos.
Não há menções diretas quanto à história e autoria das ilustrações no Theatrum, entretanto, a maioria dos desenhos é atribuída a Albert Eckhout, por semelhanças a seu estilo. Nesse sentido, haveria participação mais modesta de Frans Post na formulação dessas ilustrações.[106][107]
Historia Naturalis Brasiliae (História Natural do Brasil)
De autoria do médico Guilherme Piso (1610-1678) e do naturalista George Marcgraf (1610-1644), a Historia Naturalis Brasiliae (História Natural do Brasil)[108] foi um livro publicado originalmente em 1648. Amplamente ilustrado por gravuras (algumas feitas tendo como referência imagens presentes no Theatrum Rerum Naturalium Brasiliae), versa principalmente sobre a medicina tropical, zoologia e botânica de espécies nativas. É dividido em duas partes, sendo a primeira dedicada à medicina tropical e a segunda, composta por oito livros, destinada, sobretudo, a descrições da fauna e flora brasileira encontrada.[96][106][107]
Ao todo, 28 espécimens de crustáceos (de 24 espécies diferentes, sendo 21 de decápodes, uma de stomatopoda e duas de cracas) foram brevemente descritos e referenciados por seus nomes nativos. Destes, 18 foram ilustrados, havendo algumas incongruências entre os textos e as gravuras, dificultando a identificação das espécies — especula-se que isso tenha ocorrido por conta da edição realizada por Laet.[96]
A seguir, seguem as páginas do capítulo XIX da segunda parte da obra, constando as descrições (em latim) e ilustrações de todos os crustáceos referenciados na Historia Naturalis Brasiliae.[108]
Historia Naturalis Brasiliae: Crustacei Pisces (Crustáceos), capítulo XIX, segunda parte.
Página 182 (cap. XIX, segunda parte) de uma versão colorida de 1648.
Página 183 (cap. XIX, segunda parte) de uma versão colorida de 1648.
Página 184 (cap. XIX, segunda parte) de uma versão colorida de 1648.
Página 185 (cap. XIX, segunda parte) de uma versão colorida de 1648.
Página 186 (cap. XIX, segunda parte) de uma versão colorida de 1648.
Página 187 (cap. XIX, segunda parte) de uma versão colorida de 1648.
Página 188 (cap. XIX, segunda parte) de uma versão colorida de 1648.
Carolus Linnaeus nasceu em 23 de maio de 1707 em Raashult, na Suécia. Ele é responsável pela criação do Systema Naturae, trabalho de grande importância para o desenvolvimento da classificação taxonômica atualmente utilizada.
Ao longo da vida, ele formou duas grandes coleções de objetos relacionados à história natural, comprando itens de diversos viajantes que retornavam da Índia, América e África. Juntas, essas coleções são conhecidas como o tesauro de Seba (também conhecido como locupletissimi rerum naturalium thesauri). Dentre as diversas espécies descritas, encontram-se crustáceosdecápodes do Brasil.
Seba não utilizava o sistema de classificação de Linnaeus, apesar deste já existir na época. Para duas espécies, ele adotou a nomenclatura de Marcgraf, utilizando erroneamente o termo “uca una”, que se refere a caranguejos de mangue, para descrever caranguejos-violinistas, o que levou ao uso do termo Uca, de forma genérica, para se referir a esse grupo.
Sua ilustração de “Uka una” foi a base para a descrição de Cancer vocans major por Herbst, em 1782, posteriormente classificada como pertencente ao gênero Uca Leach, em 1814. Em 1897, para corrigir essa classificação, Rathbun e Ortmann estabeleceram, respectivamente, os gêneros Ucides e Oedipleura para classificar caranguejos de mangue, e mantiveram o uso do nome Uca Leach para os caranguejos-violinistas.
↑Wolfe, J. M., Breinholt, J. W., Crandall, K. A., Lemmon, A. R., Lemmon, E. M., Timm, L. E., ... & Bracken-Grissom, H. D. (2019). A phylogenomic framework, evolutionary timeline and genomic resources for comparative studies of decapod crustaceans. Proceedings of the Royal Society B, 286(1901), 20190079.
↑ abcdefghijklmnopqrstuvRuppert, E. E., Fox, R. S., & Barnes, R. D. (2005). Zoologia dos invertebrados: uma abordagem funcional-evolutiva. In Zoologia dos invertebrados: uma abordagem funcional-evolutiva.
↑ Chace, F. A., Jr., 1951. The number of species of decapod and stomatopod Crustacea. Journal of the Washington Academy of Sciences, 41(11): 370–372
↑ Martin, J. W. & G. E. Davis, 2006. Historical trends in crustacean systematics. Crustaceana, 79(11): 1347–1368
↑ DE GRAVE, Sammy et al. A classification of living and fossil genera of decapod crustaceans. Raffles Bulletin of Zoology, 2009.
↑ Martin, J. W. & G. E. Davis, 2001. An updated classification of the Recent Crustacea. Natural History Museum of Los Angeles County, Science Series, 39: 1–124.
↑ Tsang, L. M., et al. "Phylogeny of Decapoda using two nuclear protein-coding genes: origin and evolution of the Reptantia." Molecular Phylogenetics and Evolution 48.1 (2008): 359-368.
↑ Hagerman, Lars, and Anna Szaniawska. "Behaviour, tolerance and anaerobic metabolism under hypoxia in the brackish-water shrimp Crangon crangon." Marine Ecology Progress Series (1986).
↑ Pinn, Eunice H., and Alan D. Ansell. "The effect of particle size on the burying ability of the brown shrimp Crangon crangon." Journal of the Marine Biological Association of the United Kingdom 73.2 (1993): 365-377.
↑ Dyer, M. F., and R. F. Uglow. "Heart and scaphognathite beat behaviour in laboratory-held Crangon crangon (L.)." Journal of Experimental Marine Biology and Ecology 32.3 (1978): 209-218.
↑ Ogburn, Matthew B., et al. "Endogenous swimming activity rhythms of postlarvae and juveniles of the penaeid shrimp Farfantepenaeus aztecus, Farfantepenaeus duorarum, and Litopenaeus setiferus." Journal of experimental marine biology and ecology 440 (2013): 149-155.
↑ Chan, Tin-Yam, et al. "Phylogenetic relationships among the genera of the Penaeidae (Crustacea: Decapoda) revealed by mitochondrial 16S rRNA gene sequences." (2008).
↑ Cunha, Andressa Maria, et al. "Morphological and molecular analyses support the amphi-Atlantic distribution and taxonomic status of the snapping shrimp Alpheus intrinsecus Spence Bate, 1888 (Crustacea: Decapoda: Alpheidae)." Zootaxa4303.4 (2017): 573-589.
↑ AnKer, Arthur. "A worldwide review of stygobiotic and stygophilic shrimps of the family Alpheidae (Crustacea, Decapoda, Caridea)." Subterranean Biology 6 (2008): 1-16.
↑ Ritzmann, Roy E. "Mechanisms for the snapping behavior of two alpheid shrimp, Alpheus californiensis andAlpheus heterochelis." Journal of Comparative Physiology A: Neuroethology, Sensory, Neural, and Behavioral Physiology95.3 (1974): 217-236.
↑ Koukouvinis, Phoevos, Christoph Bruecker, and Manolis Gavaises. "Unveiling the physical mechanism behind pistol shrimp cavitation." Scientific reports 7.1 (2017): 13994.
↑ Omori, Makoto. "The biology of pelagic shrimps in the ocean." Advances in marine biology. Vol. 12. Academic Press, 1975. 233-324.
↑ Herring, Peter J. "Bioluminescence in the Crustacea." Journal of crustacean biology 5.4 (1985): 557-573.
↑ Herring, Peter J. "Bioluminescence in decapod crustacea." Journal of the Marine Biological Association of the United Kingdom 56.4 (1976): 1029-1047.
↑ Ramos-Porto, Marilena, et al. "Registro da família Eiconaxiidae Sakai & Ohta, 2005 (Crustacea: Decapoda: Thalassinidea) no Brasil, com redescrição de Eiconaxius carinatus (Bouvier 1925)." Biota Neotropica 8.3 (2008).
↑ Dalosto, Marcelo Marchet. "COMPORTAMENTO AGONÍSTICO, DESLOCAMENTO E PADRÕES DE ATIVIDADE DE LAGOSTINS NEOTROPICAIS (DECAPODA: PARASTACIDAE)." (2012).
↑ Stamhuis, EIZE J., and JOHN J. Videler. "Burrow ventilation in the tube-dwelling shrimp Callianassa subterranea (Decapoda: thalassinidea). I. Morphology and motion of the pleopods, uropods and telson." Journal of experimental biology 201.14 (1998): 2151-2158.
↑ Farmer, A. S. "The functional morphology of the mouthparts and pereiopods of Nephrops norvegicus (L.)(Decapoda: Nephropidae)." Journal of Natural History 8.2 (1974): 121-142.
↑ Smolowitz, Ronald Joel, LCDR NOAA Corps, and Northwest Fisheries Center. "Lobster, Homarus americanus, trap design and ghost fishing." Marine Fisheries Review 40.5-6 (1978): 2-8.
↑ Polovina, Jeffrey J., et al. "The role of benthic habitat, oceanography, and fishing on the population dynamics of the spiny lobster, Panulirus marginatus (Decapoda, Palinuridae), in the Hawaiian Archipelago." Crustaceana (1995): 203-212.
↑ Phillips, Bruce F., ed. Lobsters: biology, management, aquaculture and fisheries. No. 595.384 L6. Southern Gate, UK: Wiley-Blackwell, 2013.
↑ Rabalais, Nancy N., Scott A. Holt, and R. Warren Flint. "Mud shrimps (Crustacea, Decapoda, Thalassinidea) of the northwestern Gulf of Mexico." Bulletin of Marine Science 31.1 (1981): 96-115.
↑ Martin, Joel W., and Lawrence G. Abele. "External morphology of the genus Aegla (Crustacea, Anomura, Aeglidae)." Smithsonian Contributions to Zoology (1988).
↑ Pereira, Pedro Henrique Cipresso, Jairo Zancaner Junior, and Giuliano Buzá Jacobucci. "Ocupação de conchas e utilização de microambientes por caranguejos ermitões (Decapoda, Anomura) na Praia da Fortaleza, Ubatuba, São Paulo." Biotemas 22.2 (2009): 65-75.
↑ Turra, Alexander. "Comportamento, ecologia e reprodução de caranguejos ermitões (Crustacea, Decapoda, Anomura) no Sudeste brasileiro." (2003).
↑ Bertness, Mark D. “The Influence of Shell-Type on Hermit Crab Growth Rate and Clutch Size (Decapoda, Anomura).” Crustaceana, vol. 40, no. 2, 1981, pp. 197–205. JSTOR, www.jstor.org/stable/20103597.
↑ Leite, F. P. P., A. Turra, and S. M. Gandolfi. "Hermit crabs (Crustacea: Decapoda: Anomura), gastropod shells and environmental structure: their relationship in southeastern Brazil." Journal of Natural History 32.10-11 (1998): 1599-1608.
↑Cansi, Edison Rogério. "Comportamento de escape de Emerita brasiliensis (Crustacea, Anomura Hippidae); Schmitt, 1935." (2007).
↑García-Guerrero, Marcelo U., and Michel E. Hendrickx. "Embryology of decapod crustaceans, II: gross embryonic development of Petrolisthes robsonae Glassell, 1945 and Petrolisthes armatus (Gibbes, 1850)(Decapoda, Anomura, Porcellanidae)." Crustaceana (2005): 1089-1097.
↑ McLay, Colin Lindsay. Brachyura and crab-like Anomura of New Zealand. University of Auckland Marine Laboratory, 1988.
↑ Spirito, Carl P. "An analysis of swimming behavior in the portunid crab Callinectes sapidus." Marine & Freshwater Behaviour & Phy 1.1-4 (1972): 261-276.
↑ White, A. Quinton, and Carl P. Spirito. "Anatomy and physiology of the swimming leg musculature in the blue crab, Callinectes sapidus." Marine & Freshwater Behaviour & Phy 2.1-4 (1973): 141-153.
↑ Guinot, Daniēle, Dominique Doumenc, and Charles C. Chintiroglou. "A review of the carrying behaviour in brachyuran crabs, with additional information on the symbioses with sea anemones." Raffles Bulletin of Zoology 43 (1995): 377-416.
↑ Laughlin, Roger A. "Feeding habits of the blue crab, Callinectes sapidus Rathbun, in the Apalachicola estuary, Florida." Bulletin of Marine Science 32.4 (1982): 807-822.
↑ Mistiaen, Johan A., Ivar E. Strand, and Douglas Lipton. "Effects of environmental stress on blue crab (Callinectes sapidus) harvests in Chesapeake Bay tributaries." Estuaries26.2 (2003): 316-322..
↑ Johnson, David F., et al. "Wind stress and cycles in Dungeness crab (Cancer magister) catch off California, Oregon, and Washington." Canadian Journal of Fisheries and Aquatic Sciences 43.4 (1986): 838-845.
↑OLIVEIRA, Alexandre Ricardo de et al. Dieta natural do siri-azul Callinectes sapidus (Decapoda, Portunidae) na região estuarina da Lagoa dos Patos, Rio Grande, Rio Grande do Sul, Brasil. 2006.
↑ BRANCO, Joaquim Olinto; VERANI, José Roberto. Dinâmica da alimentação natural de Callinectes danae Smith (Decapoda, Portunidae) na Lagoa da Conceição, Florianópolis, Santa Catarina, Brasil. Revista Brasileira de Zoologia, v. 14, n. 4, p. 1003-1018, 1997
↑PATWARDHAN, S. S. On the structure and mechanism of the gastric mill in Decapoda. Proceedings: Plant Sciences, v. 1, n. 5, p. 183-196, 1934
↑Barker, P. L., & Gibson, R. (1977). Observations on the feeding mechanism, structure of the gut, and digestive physiology of the european lobster Homarus gammarus (L.) (Decapoda: Nephropidae). Journal of Experimental Marine Biology and Ecology, 26(3), 297–324. doi:10.1016/0022-0981(77)90089-2
↑ GOODMAN, Saul H.; CAVEY, Michael J. Organization of a phyllobranchiate gill from the green shore crab Carcinus maenas (Crustacea, Decapoda). Cell and Tissue Research, v. 260, n. 3, p. 495-505, 1990.
↑ abcdef Hobbs, H. H., & Lodge, D. M. (2010). Decapoda. Ecology and Classification of North American Freshwater Invertebrates, 901–967. doi:10.1016/b978-0-12-374855-3.00022-4
↑BAUER, RAYMOND T. Antifouling adaptations of marine shrimp (Decapoda: Caridea): gill cleaning mechanisms and grooming of brooded embryos. Zoological Journal of the Linnean Society, v. 65, n. 4, p. 281-303, 1979.
↑ REGNAULT, M. (1987). NITROGEN EXCRETION IN MARINE AND FRESH-WATER CRUSTACEA. Biological Reviews, 62(1), 1–24. doi:10.1111/j.1469-185x.1987.tb00623.x
↑ Smith, G., & Naylor, E. (2009). The neurosecretory system of the eyestalk of Carcinus maenas (Crustacea: Decapoda). Journal of Zoology, 166(3), 313–321. doi:10.1111/j.1469-7998.1972.tb03101.x
↑ McNamara, J. C. (1980). Ultrastructure of the chromatophores of Palaemon affinis Heilprin (crustacea: decapoda). The structural basis of pigment migration. Journal of Experimental Marine Biology and Ecology, 46(2), 219–229. doi:10.1016/0022-0981(80)90032-5
↑ ab SOKOLOWICZ, Carolina C.; AYRES-PERES, Luciane; SANTOS, Sandro. Diel activity and digestion time of Aegla longirostri (Crustacea, Decapoda, Anomura). Iheringia. Série Zoologia, v. 97, n. 3, p. 235-238, 2007.
↑ ab SANTOS, Marcel José Martins dos; PINHEIRO, Marcelo Antonio Amaro. Ablação ocular no camarão Macrobrachium rosenbergii (De Man)(Crustacea, Decapoda, Palaemonidae): Efeitos sobre a reprodução, pigmentação epidérmica e atividade alimentar. Revista Brasileira de Zoologia, p. 667-680, 2000.
↑OCAÑA, Frank A.; ORTIZ, Manuel; APÍN, Yanet. Nuevos registros de crustáceos marinos (Amphipoda, Isopoda, Decapoda) de aguas cubanas/New records of marine crustacean (Amphipoda, Isopoda, Decapoda) from Cuban waters. Revista de Investigaciones Marinas, v. 30, n. 3, p. 245-248, 2011.
↑Freeman, J. A., & Bartell, C. K. (1975). Characterization of the molt cycle and its hormonal control in Palaemonetes pugio (decapoda, caridea). General and Comparative Endocrinology, 25(4), 517–528. doi:10.1016/0016-6480(75)90162-8
↑ abcdefghi BRUSCA, Richard C.; BRUSCA, Gary J. Invertebrados. Madrid: McGraw-Hill, 2005.
↑ DA COSTA¹, Rogério Caetano et al. CHAVE ILUSTRADA PARA IDENTIFICAÇÃO DOS CAMARÕES DENDROBRANCHIATA DO LITORAL NORTE DO ESTADO DE SÃO PAULO, BRASIL. 2003.
↑ Baldwin, J. D., Bass, A. L., Bowen, B. W., and Clark, W. H., Jr. (1998). Molecular phylogeny and biogeography of the marine shrimp Penaeus. Mol. Phylogenet. Evol. 10:399–407
↑FAO, 2008. FAO fishery statistical collections – global capture production online query
↑Pérez Farfante, I., Kensley, B., 1997. Penaeoid and Sergestoid Shrimps and Prawns of the World. Editions du Muséum National d’Histoire Naturelle, Paris.
↑ CHACUR, M. M.; NEGREIROS-FRANSOZO, M. L. Biological aspects of the spine-shrimp Exhippolysmata oplophoroides (Holthuis, 1948)(Crustacea, Caridea, Hippolytidae). Revista brasileira de Biologia, v. 59, n. 1, p. 173-181, 1999.
↑ WICKSTEN, Mary K.; HENDRICKX, Michel E. An updated checklist of benthic marine and brackish water shrimps (Decapoda: Penaeoidea, Stenopodidea, Caridea) from the Eastern Tropical Pacific. Contributions to the study of east Pacific crustaceans, v. 2, p. 49-76, 2003.
↑ De Grave, S., Cai, Y., & Anker, A. (2007). Global diversity of shrimps (Crustacea: Decapoda: Caridea) in freshwater. Hydrobiologia, 595(1), 287–293. doi:10.1007/s10750-007-9024-2
↑ KUMAR, JS Yogesh; RAGHUNATHAN, C.; VENKATARAMAN, K. Research Article A report on some symbiotic shrimps (Crustacea: Decapoda) from the Andaman and Nicobar Islands, India.
↑ Steneck R.S., Hughes T.P., Cinner J.E., Adger W.N., Arnold S.N., Berkes F., Boudreau S.A., Brown K., Folke C., Gunderson L.,Olsson P., Scheffer M., Stephenson E., Walker B., Wilson J., Worm B. 2011. Creation of a gilded trap by the high economic value of the Maine Lobster Fishery. Conserv. Biol. 25:904–912.
↑ RICHMAN, Nadia I. et al. Multiple drivers of decline in the global status of freshwater crayfish (Decapoda: Astacidea). Philosophical Transactions of the Royal Society B: Biological Sciences, v. 370, n. 1662, p. 20140060, 2015.
↑ DWORSCHAK, Peter C. Global diversity in the Thalassinidea (Decapoda). Journal of Crustacean Biology, v. 20, n. 5, p. 238-245, 2000.
↑ Yeo, D. C. J., Ng, P. K. L., Cumberlidge, N., Magalhães, C., Daniels, S. R., & Campos, M. R. (n.d.). Global diversity of crabs (Crustacea: Decapoda: Brachyura) in freshwater. Freshwater Animal Diversity Assessment, 275–286. doi:10.1007/978-1-4020-8259-7_30
↑ RATHBUN, Mary J. The brachyura and macrura of Porto Rico. Bulletin of the United States Fish Commission, 1900.
↑ HAZLETT, Brian A. The behavioral ecology of hermit crabs. Annual Review of Ecology and Systematics, v. 12, n. 1, p. 1-22, 1981.
↑ abcdefghijklmnoVogt, Günter. “Abbreviation of Larval Development and Extension of Brood Care as Key Features of the Evolution of Freshwater Decapoda.” Biological Reviews, vol. 88, no. 1, 2012, pp. 81–116., doi:10.1111/j.1469-185x.2012.00241.x.
↑Thiel, M. Cuidado parental extendido en crustáceos – conocimiento actual. Revista Chilena de Historia Natural, 76: 205-218, 2003.
↑ Hepp LU, Fornel R, Restello RM, Trevisan A, Santos S 2012. Intraspecific morphological variation in a freshwater crustacean Aegla plana in southern Brazil: effects of geographical isolation on carapace shape. Journal of Crustacean Biology 32: 511–518..
↑ abSL de Siqueira Bueno, RM Shimizu, JCB Moraes. A global overview of the conservation of freshwater decapod crustaceans, 23-64, 2016. 13, 2016.
↑ Martin, Joel W. and Abele, Lawrence G. 1988. "External morphology of the genus Aegla (Crustacea, Anomura, Aeglidae)." Smithsonian Contributions to Zoology. 1–46. https://doi.org/10.5479/si.00810282.453.
↑Burkenroad, M. D. (1963). The evolution of the Eucarida (Crustacea, Eumalacostraca), in relation to the fossil record. Tulane Studies in Geology. 2: 3- 17.
↑Rabalais, N.N., and R.H. Gore (1985) Abbreviated development in decapods. Pp. 67–126 in Crustacean Issues 2: Larval Growth, A.M. Wenner, ed., Volume 2.
↑ abAnger, K. 2001. The Biology of Decapod Crustacean Larvae. Crustaceans Issues, 14. Lisse/ Abingdon/ Exton (PA)/ Tokyo, A.A. Balkema Publishers., p. 419.
↑Lizardo-Daudt, H. M. & Bond-Buckup, G. (2003) Morphological aspects of the embryonic development of Aegla platensis (Decapoda, Aeglidae). Crustaceana, 76(1), 13–25.
↑ abcdefghijAnger, K (2006): Contributions of larval biology to crustacean research: a review, Invertebrate Reproduction & Development, 49:3, 175-205.
↑March JG, Benstead JP, Pringle CM, Scatena FN. 1998. Migratory drift of larval freshwater shrimps in two tropical streams, Puerto Rico. Freshwater Biol. 40:261–273.
↑Anger K, Hayd L, Knott J, Nettelmann U (2009) Patterns oflarval growth and chemical composition in the AmazonRiver prawn, Macrobrachium amazonicum.Aquaculture287:341– 348.
↑Rocha, S.S.; Shimizu, R.M. and Bueno, S.L.S. 2010. Reproductive biology in females of Aegla strinatii (Decapoda: Anomura: Aeglidae). Journal of Crustacean Biology, 30: 589–596.
↑Cohen, F.P.A.; B.F. Takano; R.M. Shimizu & S.L.S. Bueno. 2011. Life cycle and population structure of Aegla paulensis (Decapoda: Anomura: Aeglidae). Journal of Crustacean Biology 31 (3): 389-395. doi: 10.1651/10-3415.1.
↑Bond-Buckup, G.; Jara, C.J.; Pérez-Losada, M.; Buckup, L.; Crandall, K.A. 2008. Global diversity of crabs (Aeglidae: Anomura: Decapoda) in freshwater. Hydrobiologia 595: 267-273.
↑Crandall K. A., and Buhay J. E.. 2008. Global diversity of crayfish (Astacidae, Cambaridae, and Parastacidae-Decapoda) in freshwater. Hydrobiologia 595: 295-301.
↑De Grave, S; Pentche , N.D.; Ahyong, S.T.; Chan, T.; Crandall, K.A.; Dworschak, P.C.; Felder, D.L.; Feldman, R.M.; Fransen, C.H.J.M.; Goulding, L.Y.D.; Lemaitre, R.; Low, M.E.Y.; Martin, J.W.; Ng, P.K.L.; Schweitzer, C.E.; Tan, S.H.; Tshudy, D. and Wetzer, R. 2009. A classi cation of living and fossil genera of decapod crustaceans. Ra es Bulletin of Zoology, Suppl. 21: 1–109.
↑Cumberlidge N, Ng PKL, Yeo DCJ, Magalhães C, Campos MR, Alvarez F, Naruse T, Daniels SR, Esser LJ, Collen B, Attipoe FYK, Ba FJC, Darwall W, McIvor A, Ram M. (2009) Freshwater crabs and the biodiversity crisis: importance, threats, status, and conservation challenges. Biological Conservation 142: 1665–1673. doi: 10.1016/j.biocon.2009.02.038
↑K. von Rintelen, T.J. Page, Y. Cai, K. Roe, B. Stelbrink, B.R. Kuhajda, T.M. Iliffe, J. Hughes, T. von Rintelen. Drawn to the dark side: a molecular phylogeny of freshwater shrimps (Crustacea: Decapoda: Caridea: Atyidae) reveals frequent cave invasions and challenges current taxonomic hypotheses. Mol Phylogenet Evol, 63 (2012), pp. 82-96.
↑Hansen JI, Mustafa T, Depledge M (1992) Mechanisms of copper toxicity in the shore crab, Carcinus maenas I. Effects on Na,K-ATPase activity, haemolymph electrolyte concentrations and tissue water contents. Mar Biol 114:253-257.
↑ abcdefghijklmnopqrstuvwxyTavares, M. S.: Toward the history of pré-Linnean carcinology in Brazil. In: Truesdale, F. (ed.): History of Carcinology. Crustacean Issues, v. 8, A. A. Balkema, Rotterdam. pp. 7-29, 1993.
↑Leitão, C. M.: História das expedições científicas no Brasil. São Paulo: Editora Nacional. 1941.
↑Barbosa, Pe. A. L.: Pequeno vocabulário tupi-português. Rio de Janeiro, Livraria São José. 1951.
↑ abHolthuis, L. B.: The subfamily Palaemoninae. A General Revision of the Palaemonidae (Crustacea, Decapoda, Natantia) of the Americas. II. Occ. Pap. Allan Hancock Found. 12: 1-396.
↑Pires, F. P.: Viagem à Terra do Brasil: Jean de Léry: entre a medievalidade e a modernidade. Revista Caminhando, v. 8, n. 1, pp. 89-112, 2003.
↑ abSilva, W. C. L.: A invenção de um olhar: Jean de Léry e os tupinambás. História Social, Campinas, n. 8/9, pp. 71-104, 2001/2002.
↑Tavares, M. S. & Albuquerque, E. F.: Levantamento taxonômico preliminar de Brachyura (Crustacea: Decapoda) da Lagoa de Itaipu, Rio de Janeiro, Brasil. Atlântica. 11(1), pp. 101-108, 1989.
↑Léry, J. de: Viagem à Terra do Brasil. Tradução integral e notas de Sérgio Milliet. Editora Biblioteca do Exército, 1961.
↑ abcdefghiAlbertin, P. J.: Arte e ciência no Brasil holandês: Theatri Rerum Naturalium Brasilae: um estudo dos desenhos. Revista brasileira de zoologia, São Paulo, 3 (5), pp. 249-326, 1985.
↑ abcdefgGesteira, H. M.: O Recife holandês: história natural e colonização neerlandesa (1624-1654). Revista da SBHC, Rio de Janeiro, v. 2, n. 1, pp. 6-21, 2004.
↑ abMarcgraf, G. & Piso, G.: Historia Naturalis Brasiliae... in qua non tantum plantae et animalia, sed et indigenarum morbi, ingenia et mores describuntur et iconibus supra quingentas illustrantur. Lugdun. Batavorum, apud Franciscus Hackium et Amstelodami apud Lud. Elzevirium. Joannes de Laet (org.), 1648, pp. 182-188. Digitalizado e disponipilizado por: Peter H. Raven Library/Missouri Botanical Garden.

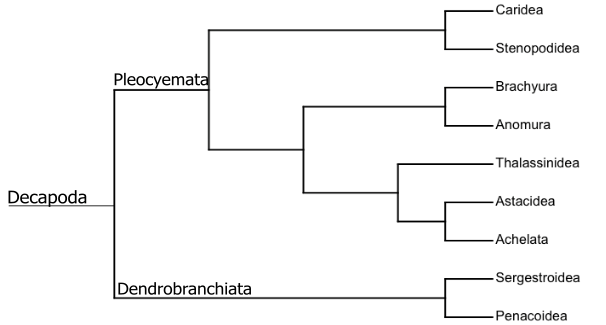

_(23051697572).jpg)
.jpeg)






.jpg)










 Página 182 (cap. XIX, segunda parte) de uma versão colorida de 1648.
Página 182 (cap. XIX, segunda parte) de uma versão colorida de 1648. Página 183 (cap. XIX, segunda parte) de uma versão colorida de 1648.
Página 183 (cap. XIX, segunda parte) de uma versão colorida de 1648. Página 184 (cap. XIX, segunda parte) de uma versão colorida de 1648.
Página 184 (cap. XIX, segunda parte) de uma versão colorida de 1648. Página 185 (cap. XIX, segunda parte) de uma versão colorida de 1648.
Página 185 (cap. XIX, segunda parte) de uma versão colorida de 1648. Página 186 (cap. XIX, segunda parte) de uma versão colorida de 1648.
Página 186 (cap. XIX, segunda parte) de uma versão colorida de 1648. Página 187 (cap. XIX, segunda parte) de uma versão colorida de 1648.
Página 187 (cap. XIX, segunda parte) de uma versão colorida de 1648. Página 188 (cap. XIX, segunda parte) de uma versão colorida de 1648.
Página 188 (cap. XIX, segunda parte) de uma versão colorida de 1648.,_segunda_parte,_cap._XIX,_p._182.jpg)
,_segunda_parte,_cap._XIX,_p._183.jpg)
,_segunda_parte,_cap._XIX,_p._184.jpg)
,_segunda_parte,_cap._XIX,_p._185.jpg)
,_segunda_parte,_cap._XIX,_p._186.jpg)
,_segunda_parte,_cap._XIX,_p._187.jpg)
,_segunda_parte,_cap._XIX,_p._188.jpg)